
tags: parrots, Psittaciformes, evolution, molecular phylogeny, ornithology, Neornithes

Red-crowned Amazon parrot, Amazona viridigenalis, at Elizabeth Street Parrotry, Brownsville, Texas.
Image: Joseph Kennedy, 7 April 2008 [larger view].
Nikon D200, Kowa 883 telescope TSN-PZ camera eyepiece 1/750s f/8.0 at 1000.0mm iso400.
![]()
One of the most contentious issues among scientists who study the evolution of birds is identifying precisely when the modern birds (Neornithes) first appeared. This is due to conflicts between the fossil record and molecular dating methodologies. For example, fossils support a Tertiary radiation whereas molecular dating methodologies suggest that the birds radiated in the early Cretaceous. But there is another way to address this discrepancy. Because the evolution of parrots and cockatoos reflects the evolution of Aves themselves, studying the psittaciformes offers compelling insights into this mystery. Further, because psittaciformes generally are not migratory and because they tend to occupy discrete ranges, their ancient patterns of diversification are easier to discern than for many other taxonomic orders of birds that have dispersed widely.
To this end, an international team of researchers, headed by Timothy Wright, a professor at New Mexico State University and a researcher at the Smithsonian Institution in Washington DC, collaborated to study the early evolution of the parrots. To do this work, they sampled individual representatives from 69 of the 82 extant genera of parrots and eight additional avian orders that have been idnetified as outgroups, for a total of 77 avian taxa. Outgroups are defined as those taxa that possess a character, such as a hooked beak or zygodactyl toes, that is likely to have been ancestral to the parrots.
Wright and his colleagues chose the Falconiformes (Falcons), Passeriformes (songbirds), Columbiformes (pigeons and doves), Cuculiformes (cuckoos), Piciformes (woodpeckers), Coraciiformes (the "near passerines", such as kingfishers, etc), Strigiformes (owls), and Coliiformes (mousebirds) as outgroups because each was considered an ally of the parrots at one time or has been identified as a sister group in recent molecular phylogenetic work [for example, see here].
The team sequenced and analyzed 3,915 basepairs (bp) of DNA per individual, consisting of 1,668 bp from mitochondrial DNA (mtDNA) that encodes two genes, and another 2,247 bp from nuclear (chromosomal) introns. Using these data, they used Maximum Parsimony analysis to recover this cladogram that describes the relationships between the 69 parrot genera and the eight outgroups (figure 1);
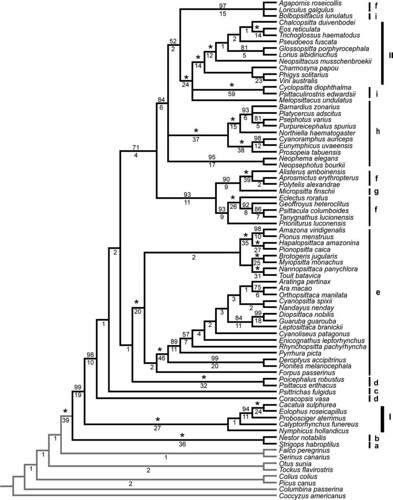
FIG. 1. -- Cladogram of relationships among representatives of 69 parrot genera and 8 outgroup taxa based on a strict consensus of 4 trees obtained from combined parsimony (MP) analysis of the 4 sequence partitions (COI + ND2, TROP, TGFB2, and RDPSN) with coded gaps. MP bootstrap support values for each node are shown above the branches with values of 100% indicated by asterisks; Bremer decay index values are shown below. The classifications of Rowley (1997) and Collar (1997) are indicated by bars on the right: I) family Cacatuidae, II) subfamily Loriinae, and, within the subfamily Psittacinae, the tribes a) Strigopini, b) Nestorini, c) Psittrichadini, d) Psittacini e) Arini, f) Psittaculini, g) Micropsittini, h) Platycercini, and i) Cyclopsittacini [larger view].
DOI: 10.1093/molbev/msn160.
The team then used Bayesian analysis to reconstruct the evolutionary history of the parrots and recovered this phylogram, which describes very similar relationships to those in the above cladogram (figure 2);
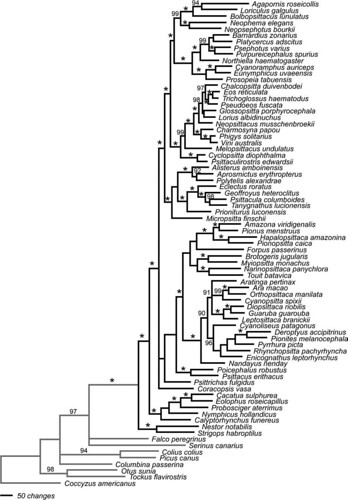
FIG. 2. -- Phylogram of relationships among representatives of 69 parrot genera and 8 outgroup taxa based on Bayesian analysis of the 4 sequence partitions (COI + ND2, TROP, TGFB2, and RDPSN) and a fifth partition consisting of coded gaps from the 4 sequence partitions. Bayesian posterior probabilities 0.90 are indicated as percentages above the branches, values of 1.0 are indicated with an asterisk. The scale bar indicated the number of changes (base substitutions or changes in state of indel characters) [larger view].
DOI: 10.1093/molbev/msn160.
The team then estimated divergence times using both two different analytic methods that relied upon incorporation of one of the two contrasting calibration dates (either based on fossil data and molecular data) (figure 3);
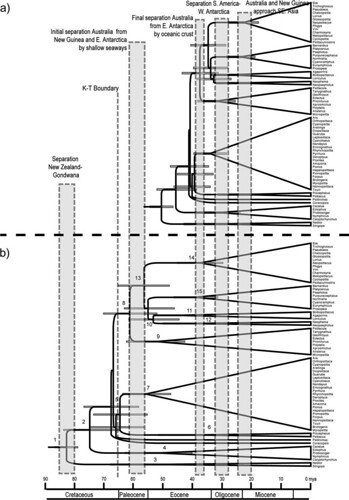
FIG. 3. -- Chronograms showing divergence times among parrot genera based a Bayesian relaxed-clock approach, with 2 alternative dates for the basal divergence between the New Zealand endemics Nestor and Strigops and the remaining psittaciforms: (a) node dated to 50 MYA, based on the hypothesized divergence between modern psittaciforms and fossils dated to the Lower Eocene; and (b) node dated to 82 MYA, based on the split between New Zealand and Gondwana. Uncertainty in the timing of these alternative divergences is incorporated with both calibration estimates representing means of normal distributions with SD 5 2; error bars on the nodes illustrate the 95% HPD of node ages. Shaded vertical bars indicate timing of major geological events that may have contributed to diversification through vicariance: separation of New Zealand from Gondwana, 80-85 MYA; the K/T boundary, 65 MYA; the initial separation of Australia from East Antarctica, 58-61 MYA; the final separation of Australia from East Antarctica, 36-41 MYA; the final separation of South America from West Antarctica, 28-32 MYA; and Australia and New Guinea approach southeast Asia, 20-24 MYA [larger view].
DOI: 10.1093/molbev/msn160.
When the team used the molecular calibration date of 82 million years ago (MYA), they found that the evolutionary split between the cockatoos and the remaining psittaciforms occurred prior to the K/T boundary at 65 MYA. Subsequent parrot lineage radiations then occurred between 60 and 65 MYA, which corresponds to the initial separation of Australia and New Guinea from East Antarctica. According to this estimate, diversification into the major parrot subclades was complete by 30 MYA, by which point Australia, Antarctica, and South America were completely separated.
On the other hand, when the team instead used 50 MYA (fossil data) to calibrate the diversification in basal parrot lineages, they found that the split between the cockatoos and the remaining psittaciforms occurred 45.04 MYA -- just prior to the time Australia is thought to have separated from Antarctica. Radiation of Neotropical parrot lineages is estimated to have occurred starting 33 MYA, a period roughly coincident with the separation of South America from West Antarctica.
These results support the hypothesis that Australasia was probably where the parrots originated. This hypothesis is based in the high number of genera endemic to the region, several of which are rooted at or near the base of the psittaciform tree. Further, the basal split between the New Zealand taxa, Nestor and Strigops, suggests that this origin occurred while New Zealand and Australasia were still part of the Gondwana supercontinent during the Cretaceous period. Additionally, other recently published analyses using similar methods suggest that both Passeriformes and Columbiformes originated in Gondwana during the Cretaceous.
Other interesting findings include the confirmation that the cockatoos are a monophyletic group -- as are the lories (Loriinae), whose sister groups include Melopsittacus, Cyclopsitta, and Psittaculirostris -- a finding that agrees with my data (unpublished). Unsurprisingly, these data indicate that the remaining parrots form a polyphyletic jumble, with the exception of the tropical African parrots and the Neotropical Arini, each of which were recovered as separate well-supported clades.
The team also found that their various phylogenies did not produce any consistent placement of the eight outgroups as sister to the parrots, reinforcing the idea that the parrots and cockatoos do not have any close sister relationships with other modern birds.
This research indicates that Aves appeared during the Cretaceous, and that parrots are an ancient lineage without any close evolutionary relationships. Additionally, these data also support the hypotheses that there are several separate and well-defined clades of parrots, particularly cockatoos and the lories, while the remainder of the parrots need some serious phylogenetic work to be done -- work that I would absolutely love to devote my life to completing.
Source:
T. F. Wright, E. E. Schirtzinger, T. Matsumoto, J. R. Eberhard, G. R. Graves, J. J. Sanchez, S. Capelli, H. Muller, J. Scharpegge, G. K. Chambers, R. C. Fleischer (2008). A Multilocus Molecular Phylogeny of the Parrots (Psittaciformes): Support for a Gondwanan Origin during the Cretaceous. Molecular Biology and Evolution, 25 (10), 2141-2156 DOI: 10.1093/molbev/msn160.
- Log in to post comments
informative as well as enlightening... would love to learn more, and specifically the Anodorhynchus hyacinthinus, and the other blue variants...
What parrots tell us about professional gift wrapping services.
I'm a casual reader who followed a link from slash dot. I'm missing the point. The way I read the story, theoretical analysis of the molecular data shows cockatoos branched of 60-65 MYA. Fossil data indicates 45 MYA. If the fossil data gave an older age I would assume the theoretical analysis was wrong. But when the fossil data gives a younger age, one might blame it on missing fossils. Is the fossil record sufficiently complete to clearly prohibit this interpretation? Is one data set a clear winner? -- Thanks
My God, how am I suppose to read figure 3?
Your analysis is incomplete and it shows. Do your homework next time.
'But when the fossil data gives a younger age, one might blame it on missing fossils.'
Agreed. Writer needs to RTFA before posting it...
Isn't Merle's Door a neat book? Loved it...
Thanks for the parrot info -- fascinating!
Read the larger copy of the phylogeny tree at Flickr.com by clicking on the picture. You will need to set up an account there or see if this works
http://www.flickr.com/photos/grrlscientist/3118833840/sizes/l/
1. Fossils are sometimes incorrectly dated.
2. Doing a Sherlock Holmes number (poetically speaking) on molecular gene material ain't exactly like making jello.
" To do this work, the Early Bird group examined
19 independent loci consisting of 32 kilobases of nuclear DNA sequence obtained from 169 avian species representing all major groups of birds alive today. These nuclear loci included introns (74%), exons (coding regions, or "genes") (23%), and untranslated regions (UTRs) (3%) across 15 different chromosomes (according to the chicken genome). These DNA sequence data were subjected to multiple statistical analysis that produced a robust phylogeny."
Ton of stuff going on there with huge boatloads of assumptions probably. Good work, but if some of the assumptions are wrong? ? ?
Wow, this tree had a few surprises. I'll mention a few things I find particularly interesting.
I have to add that, using this cladogram and my dislike of lumping Psittaciforms into one 'family' I have this division in mind here:
Nestoridae for Nestor and Strigops.
Cacatuidae for cockatoos and cockatiel.
Psittacidae for Psittrichas, Coracopsis, the African parrots and the American parrots.
Psittaculidae for the psittaculines, polytelitines and Micropsitta.
Loriidae for lories, fig-parrots and Melopsittacus.
Platycercidae for Australian broad-tailed parrots, Cyanoramphus, Eunymphicus and Prosopeia. This group would probably include Psittacella too.(Recent research placing it as the basalmost 'broad-tail'.
Where Agapornis, Loriculus and Bolbopsittacus go isn't clear. They're either loriids, platycercids or yet another 'family'.
- First, it seems that the bauplan of parrots inhabiting the dry interior of Australia (small bills, long pointed wings, long pointed tails, speculum as well as a nomadic lifestyle and non-seasonal breeding) has developed at least five times. Nymphicus for the cacatuids, Polytelis for the psittaculids, Neophema and Neopsephotus on one hand, and Psephotus and Northiella on two other hands for the platycercids and finally Melopsittacus for the loriids. This suggests some very strong evolutionary pressure.
- Second, the psittacid tree shows a weird distributional pattern: The most basal genus Coracopsis is Madagascan, next comes Papuan Psittrichas, then African Psittacus and Poicephalus and finally, the Neotropan assemblage. Since I'm highly sceptical of modern parrots evolving in the Cretaceous, this seems to suggest a double invasion of Africa (and by extension the Americas) via Kerguelen in the Palaeogene.
- The Neotropical parrots are highly divergent morphologically. I would have expected Rhynchopsitta to end up near Ara, for example. This suggests the strong bill of Rhynchopsitta is convergent with that of Ara, rather then homologous. It's a pity Anodorhynchus, Primolius and Ognorhynchus weren't included. It would have been very interesting to see where these would have ended up, given that we see a macaw clade (excluding Diopsittaca, which is sister to Guaruba)
- Pionopsitta caica should have been Gypopsitta caica. With that in mind, it would have been interesting to see where Pionopsitta pileata would have ended up given that it's the only currently recognised member of its genus. Also, Triclaria is sorely missed. According to parrot expert Rosemary Low, it's close to Pionopsitta.
- Lastly, Bolbopsittacus in a trichotomy with Agapornis and Loriculus is unexpected. It was previously considered related to either fig-parrots or Psittinus. However, this does result in a nice clade of small, shorttailed Afro-Asian parrots that is geographically 'clean'.
What do you think of this?
Wow! Most of this is beyond my level of understanding due to my limited background with birds, but was very interesting and challenging for me to digest.
Thanks for making me reach out.