
Just when I'm in the middle of talking about the new dinosaurs and pterosaurs that were published in January (part I, part II, part III), February is turning out to be just as interesting. Yesterday saw the debut of the new basal sauropodomorph Panphagia protos (Martinez & Alcober 2009), and you'll know from SV-POW! that Matt Wedel's long-awaited paper on pneumatic hiatuses in sauropods recently appeared in Journal of Experimental Zoology.
Today sees the appearance of another very neat study: Leon Claessens, Pat O'Connor and David Unwin's research on respiration and pneumaticity in pterosaurs (Claessens et al. 2009). Yet again here is an excellent paper that's available no-holds-barred to anyone, thanks to the wonder of open-access publishing. It joins another outstanding paper on pterosaur palaeobiology, also published in an open-access journal, and which modesty prevents me from mentioning [and, on another matter entirely, you will note that the promotional picture used above is by my co-author Mark Witton].
Anyway, those of you with good memories will recall that the conclusions of the current study were mentioned here at Tet Zoo back in September 2007. This is when the data was presented at the Munich Flugsaurier conference held in honour of Peter Wellnhofer. September 2007 is a long time ago: how come it took so long for this work to appear in print? You will note that, coincidentally, another paper on ornithodiran pneumaticity appeared not too long ago in the same journal, but I'm sure that has nothing to do with it.
Claessens et al. (2009) essentially looks at two different, but connected, details of pterosaur palaeobiology: (1) how pterosaurs breathed, and (2) how pterosaurs pneumatised their wing skeletons. On (1), a good argument is made that pterosaurs pumped their big sterna dorsoventrally like birds do, and that it was this action that drove the breathing cycle. The mobile sternum and its associated sternal ribs explain how pterosaurs could breathe effectively even when the dorsal vertebrae were fused together to form a notarium: as Claessens et al. (2009) note, the presence of elaborate sternocostapophyses [read on] in basal pterosaurs shows that a reliance on 'sternal pumping' predated the evolution of the notarium.
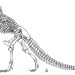
Several details of pterosaur anatomy, previously mischaracterised or misunderstood, provide further support for this model. It turns out that the sternal ribs are not the same length all the way along the sternum, but that the most posterior elements are at least twice as long as the most anterior ones. Furthermore, the sternum was not horizontal or posterodorsally sloped in the body, but posteroventrally sloped, thereby making the part of the trunk adjacent to the posterior border of the sternum by far the deepest part of the body. Sternal ribs are typically simple and rod-shaped, but those of pterosaurs possess dorsal and ventral processes that Claessens et al. (2009) term sternocostapophyses [image below, from the paper, shows sternal ribs with sternocostapophyses in Rhamphorhynchus. Anterior is to the right: the big bone at bottom right is the sternum]. They propose that these structures increased the leverage of the intercostal muscles, and thereby helped sternal movement during respiration. The sternocostapophyses therefore seem to be functional analogues of the uncinate processes present on the ribs of maniraptoran theropods, and you'll recall what uncinate processes might be for because we looked at it just the other day in connection with kiwi.
It is further hypothesised that the prepubes - mobile fan-shaped bones (unique to pterosaurs) that articulated with the pelvis and covered the posterior part of the belly - and the gastralia (the rod-like 'belly ribs' embedded within the abdominal wall) also worked together during breathing. During inspiration, downward and backward rotation of the prepubes would have increased pulmonary space: this is essentially similar to what happens in extant crocodilians, except that in their case it's the mobile pubic bones that fulfil this role.
Because this functional complex - involving the sternum and sternal ribs anteriorly and gastralia and prepubes posteriorly - is consistent across the whole of Pterosauria, Claessens et al. (2009) argue that it applied generally to all members of the group.
A rant about endothermy
There's more to this study than a model of respiration in pterosaurs: they also looked at pneumaticity. It's been clear for a while now - well, it has to everyone who actually works on dinosaurs and pterosaurs, anyway - that pterosaurs and saurischian dinosaurs exhibited both skeletal and soft-tissue pneumatisation. The presence of an air-sac system connected to the lungs shows that a bird-like flow-through system was present, and that oxygen extraction was therefore relatively high. Given that pterosaurs were active flapping fliers with huge muscles, it appears logical to conclude that a bird-like style of pulmonary ventilation helped fuel an elevated metabolism [image below shows Matt Wedel's reconstruction of the respiratory system in a diplodocid sauropod, from the pneumaticity tutorial at SV-POW!].
Many palaeontologists seem to keep quiet about ornithodiran palaeophysiology these days, which is ironic given that a large amount of data supporting endothermy has been published in recent years. All too often we see colleagues playing at being conservative, and implying or stating that ectothermy in Mesozoic archosaurs is likely or viable. Then again, maybe the conservative workers are the more vocal ones. Given the good evidence we now have for extensive pneumatisation, fibrous integumentary structures, erect gaits and erect neck carriage, and high growth rates in ornithodirans, the conclusion that these animals were endothermic appears well supported, whereas it is not possible to say the same for the ectothermy hypothesis. It's interesting that the several bits of hard evidence meant to support ectothermy in ornithodirans, like the presence of a hepatic piston diaphragm pump and an absence of respiratory turbinates, are erroneous or highly suspect.
So, after going for a long time not really caring either way about extinct archosaur physiology, I would currently argue that the evidence for endothermy is good, and that data indicates that all dinosaurs and pterosaurs were endothermic.
Pterosaurs get SASSy
Anyway, Claessens et al. (2009) were particularly interested in the distribution of pneumatic foramina within the appendicular skeletons of pterosaurs. This is mostly restricted to ornithocheiroids and azhdarchoids, and notably these are the groups that achieved giant size. The fact that the pneumatic foramina of ornithocheiroid and azhdarchoid limb bones are located in different places indicates that they evolved this appendicular pneumatisation independently.
Even more interesting is the possibility that the source of this pneumatisation in the wing elements may have been a subcutaneous air sac system (SASS): a complex system of sacs located beneath the skin. This might sound bizarre but it's present in many large modern birds, including vultures, pelicans and bustards. One pterosaur specimen - an exceptionally well-preserved chunk of wing membrane (Martill & Unwin 1989) - even preserves a layer of spongy internal tissue that, presumably, represents part of this SASS. The fact that the external cutaneous surface of the pterosaur wing was the same thing as the wing surface itself means, you'll note, that the SASS might have played a different role in pterosaurs than it does in birds. Claessens et al. (2009) speculate, for example, that differential inflation of certain regions of the SASS may have been used to control the mechanical properties of the wing. They even allow the possibility that SASS inflation could have been used in display, just as it is in some extant birds (Akester et al. 1973). We might infer that such complex control of subcutaneous wing structures would require sophisticated neural feedback. The good news is that we have good evidence from pterosaur brain anatomy that this is exactly what they had (Witmer et al. 2003).
The inevitable stuff about giant size
One of the greatest questions about pterosaurs, and one of the aspects of their palaeobiology that makes them different from other volant vertebrates, concerns their size (we'll ignore teratornithids and pelagornithids for now, sorry). Pterosaurs belonging to at least two lineages (Ornithocheiroidea and Azhdarchoidea) evolved gigantic size, and members of one of these clades (Azhdarchoidea) became substantially larger than all other fliers (including teratornithids and pelagornithids) [image below: an ornithocheirid by Mark Witton. From here].
Why pterosaurs evolved giant size is an interesting question. There is no reason to think that special atmospheric conditions, or a different global climate, or special environments, allowed them to do this: my take on the evolution of giant size has always been pretty simple... animals get big because they can. In other words, they inherit anatomical details and a physiology that allows them to grow into giants should selection favour it. I think it's increasingly clear that the weird animals of the Mesozoic were not really any weirder than the animals that came before or after. We have to get over the idea that dinosaurs and pterosaurs were freakish: that is, it seems that they hadn't evolved totally novel solutions to problems (like a unique, ontogenetically variable physiology, or a reliance on unusual atmospheric conditions, or a system of balloons, or multiple hearts, or gill slits, that allowed them to hold up and breathe through their long necks). Instead, they were just 'extreme' examples of what morphological and physiological variation could allow.
Is it possible that the extensive pneumatisation present in pterosaurs contributed to their ability to evolve giant size? Claessens et al. (2009) comment on this, but they don't actually say much (despite the title of the paper!) other than that, yes, it probably did.
Incorporating CT-scanning and x-ray data, and including inferences on pneumaticity, palaeophysiology and evolutionary trends, I'm sure you'll agree that this is a neat study that really augments our understanding of pterosaur biology. Congrats to the authors, and commiserations on the long and inexplicable delay. Remember to visit PLoS ONE and download the paper!
Refs - -
Akester, A. R., Pomeroy, D. E. & Purton, M. D. 1973. Subcutaneous air pouches in the Marabou stork (Leptoptilos crumeniferus). Journal of Zoology 170, 493-499.
Martill, D. M. & Unwin, D. M. 1989. Exceptionally well preserved pterosaur wing membrane from the Cretaceous of Brazil. Nature 340, 138-140.
Witmer, L. M., Chatterjee, S., Franzosa, J. & Rowe, T. 2003. Neuroanatomy of flying reptiles and implications for flight, posture and behaviour. Nature 425, 950-953.
More like this


Very interesting. Would a skeletal breathing pump linked to flapping explain the long necks of azhdarchids? Compare our swans: longer wings make for more efficient flight, but also mean a lower wing beat frequency. That means longer, bigger breaths, which brings the danger of hyperventilation. Swans have evolved long necks, and even longer oesophagus coiled around the inside of their bodies, to breath deep without hyperventilating.
Great post! This is why I am a tetra addict.There are many of us who don't have the academic background to easily plough through a specialized academic article, yet sneer at the poor attempts of the main stream press to cover this lost world.
This sight satisfies that itch better than any other.
Thanks for the post - I echo what thylacine posted. I love this stuff.
Derek, I assume you mean that it's the even longer trachea, not oesophagus, coiled within the body of the swan, that prevents hyperventilation. But this in any case is news to me -- do you have a reference for this? It's not obvious how passing breaths down a long trachea would prevent hyperventilation.
Excellent, fascinating material.
However I am not sure about your comment: "There is no reason to think that special atmospheric conditions, or a different global climate, or special environments, allowed them to do this"? Surely there is evidence that oxygen levels, CO2 levels and overall atmospheric density have varied over time and logically this would be a factor in determining what the physical limits of what were possible for organisms at any given period were. So while such atmospheric factors wouldnt be the cause of the large size, it would affect the limits of what is possible.
Also you state that "data indicates that all dinosaurs and pterosaurs were endothermic" - what data indicates ornithischian dinosaurs were endothermic? (I hope its not just phylogenetic bracketing). I can see some for saurischians, though even this is not without its problems. I saw some calculations a few years ago of how much fodder a large sauropod would require to be truly endothermic, and while it was possible for some, the largest sauropods seem to have well exceeded the threshold of what was possible (the calculation suggested that even with very high quality material, and non-stop 24 hour a day eating, the largest sauropods couldn't consume enough calories to be truly endothermic. Of course thats not to say they couldnt have raised their metabolism slightly and their mass then result in them being functionally, but not truly endothermic.).
The case for pterosaurs being endothermic is to me quite compelling.
Thanks again for a superb commentary. I haven't read your paper thoroughly yet but it looks excellent, and thanks for getting it published in PLOS.
Trachea. Windpipe.
Interesting idea.
Sure, but even the biggest azhdarchids would be completely viable in today's atmosphere.
Growth patterns, bone histology, way of breathing, adaptations to cursoriality and the likeâ¦
That calculation was stupid enough to assume that sauropods spent lots of time chewing like elephants. They didn't.
And how do you explain the air-sac system and the impressive growth rates?
But⦠a high temperature cannot power the muscles.
Approach the problem the other way around: The metabolism of the heart, gut, and the muscles used for just standing would on its own make a large sauropod endothermic (Paul & Leahy 1994, Paul 1998).
The only known gigantotherm is the leatherback turtle, which has lots of features not seen in sauropods (thermal insulation by shape, shell, fat layer, and the fact that the constantly working locomotor muscles are inside the shell!) and has a metabolism so sluggish it couldn't walk at a reasonable speed no matter what its anatomy.
(That is, Dermochelys is endothermic, because its body heat comes from its muscles, not from the cold sea, and it is reasonably homeothermic. It is, however, bradymetabolic and bradyaerobic, and both of the latter can be ruled out in sauropods.)
http://palaeo.jconway.co.uk/pterosaur_structure.php
John Conway had done an illustration of this point some months ago.
Endothermy relates not only to maintainance of a high and constant core body temperature, but also relates to the way in which that temperature is generated (in both birds and mammals the excess heat is effectively generated by the action of hyperactive sodium potassium pumps in the cell membranes, whose activity is necessary as both of these groups have higher porosity to these ions that usual among other vertebrates). The evolution of endothermy is effectively the evolution of leaky cell membranes. This cannot be tested effectively in a fossil taxon nor implied for one outside of the known crown groups possessing this feature. All of the features listed by Darren may indicate either elevated metabolic rates or some element of heat conservation (whether by gigantothermy, activity or insulation or some combination thereof) - however, they do not necessarily provide any evidence for how those higher core temperatures are generated or maintained. Unless you can shown intracellular metabolism is the underlying reason for elevated and constant core temperatures you cannot prove endothermy. Personally, I think it is clear that many ornithodirans had elevated metabolic rates and/or used other mechanisms to help maintain core temperatures at higher than ambient. However, this is not the same as proving endothermy, which refers to a specific set of metabolic processes.
Wow...Nemo Ramjet was right. In case you don't know what I'm talking about, Nemo wrote a short story a while back called Hatzegoptery Rape (yes, really), in which a group of time travelers bring an azdarchoid back from the Cretaceous, and it ends up attempting to make love to a giraffe, the giraffe kicking it in the chest and killing it. But the point is, Nemo's azdarchoid had the same inflatable air sacs to keep it aloft.
Anyway, now we know that pterosaurs and non-avian dinosaurs had bird-like lungs, which joins many features which show that non-avian dinosaurs, birds, and pterosaurs form a natural clade in tetrapod taxonomy (in addition to dinofuzz, hollow bones, and other features). So take that Dave Peters, and all y'alls flying lepidosaurimorphs!
High five Darren! Welcome the endothermic ornithodiran club!
But more on subject, I see a panoply of way in which sauropods could conform to this model of ornithodiran endothermy. First off, sauropods could have become secondarily ectothermic, like crocodilians did when they returned to the water. Secondly, sauropods could have had a rather low natural endothermic metabolism (think sloth), but were still able to move like full endotherms due to fermentative endothermy, gigantic endothermy, etc. My favorite explanation is the "gear-switching" model. In normal warm conditions, sauropods did not have to create much heat endothermically, instead fermentative endothermy, gigantothermy, and other mechanisms were able to provide a lot of the heat, allowing the sauropod to focus its energy on other things (growth, movement, etc.) However, when the climate got colder, the sauropod's metabolism could kick into high gear, supporting itself more by its own endothemy now than just fermentative endothermy and such. But once again, these are just theories.
I also applaud you on going against the whole "birds got as big as they could with Argentavis" and such. No. If environmental conditions suit bigger flying animals, then bigger flying animals will develop. Pterosaurs were probably better adapted for larger flight than the current two volant tetrapod groups (bats and birds), but it probably didn't help as much as many people claim. I have heard so many times that "x could not exist", or "y could not exist", x and y most often referring to Stomatosuchus and azdarchoids. But most of the time, the reason why we don't see azdarchoid-sized teratorns in the Miocene and such is because the environment provided no benefits for a giant flier vs. a smaller one. Evolution doesn't work as Napoleon Dynamite put it "because it felt like it, gosh!".
By the way, you might be interested in Unwin's book "The Pterosaurs: From Deep Time" if you don't already have it. In it, Unwin talks about how they attempted to come up with the size limits for flight, and came up with the estimate that flying creatures (esp. azdarchids) could, in theory, grow large enough to have sixty-six foot wingspans! Weird.
Well, I haven't read the paper yet -- it's downloading presently -- so I don't know what, if anything, the authors say about this, but this strikes me as odd -- do birds really have the ability to "differentially" inflate/deflate their air sacs? I have been under the impression that they could not -- there is no system of valves in the sacs or diverticula that can trap air in or out; they're either inflated or deflated. This was, as I recall, the main objection to Akersten's hypothesis that cervical air sacs provided neck support in sauropods by creating a turgid yet very lightweight means to hold their necks up -- there's simply no way to keep the air in there under high pressure separate from the remainder of the respiratory system...?
Or, by "differential," did you simply mean the dichotomous "inflated" and "deflated" states?
And for good reason. The evidence in favour of endothermy is rather untested in extant ectotherms, or just plain equivocal. Speedy bone growth was once thought to be evidence of endothermy, until it was found that both giant tortoises and crocodylians sport this style of growth (e.g. Tumarkin-Deratzian, 2007). Dinosaur growth was originally viewed as blazing fast (Erickson, 2001), but a review of the data revised that growth rate down substantially (Lehman and Woodward, 2008). Erect stance is a red herring as many crocodylomorphs had this, as well as rauisuchians and pareiasaurs (unless one wants to imply automatic endothermy to all these guys too). Same for assumptions based off of paleo-latitude (i.e. dinosaurs appear in polar strata that also housed other reptiles).
The safe bet, or the "conservative stance" in this case, is to look at this data as equivocal, since that's what scrutiny of the data has shown it to repeatedly be.
You're right, it's trachea, and I misremembered that it was the length that counted. It's the dead space in proportion to the tidal volume that counts, mixing to produce a suitable concentration of CO2 in the posterior sacs before entering the lung.
I have it from Knut Schmidt-Nielsen's How animals work, page 49 (1972), and Schmidt Nielsen in turn references Hinds and Calder, Evolution, 25 429-40 (1971)
We've come a long way from the days when pterosaurs were the victims of scientific slander, believed to have flown poorly, if at all. Now all the lines of evidence point clearly to creatures adapted to an active, flapping lifestyle, with adaptations no one had guessed at. By the way, Unwin was one of those who hedged at pterosaurian endothermy in his book, "Pterosaurs: From Deep Time". Presumably the accumulating evidence has caused him to change his mind.
"Endothermy relates not only to maintainance of a high and constant core body temperature, but also relates to the way in which that temperature is generated" (Paul Barrett)
Since when? 'Endothermy' refers only to the ability to maintain a constant high internal temperature by way of internal means and does not only refer to mammal or bird-style cell biology. cf. endothermic fish (scombroids, pelagic opah, lamnids), wasps, moths. If dinosaurs were able to maintain a high internal temperature by way of internal means, then by definition they were endothermic.
Speedy bone growth was once thought to be evidence of endothermy, until it was found that both giant tortoises and crocodylians sport this style of growth (e.g. Tumarkin-Deratzian, 2007).
Yeah, you can get tortoises and crocs to lay down fast-growing bone for a while, if you keep them warm and give them plenty to eat. But they can't do it for their entire lives or in seasonally variable environments, which is what birds and mammals can do.
Sauropod growth rates have been problematic because of different methods of retrocalculating LAGs. The Erickson et al. growth rates are probably too fast, and the Lehman and Woodward growth rates are probably too slow. The most complete set of LAGs in a single individual sauropod is in the giant mamenchisaur presented at SVP a couple of years ago by Wings et al., which grew to adult size in three decades--about twice as long as projected by the Erickson et al. curve, and about half as long as projected by the Lehman and Woodward curve. In any case, we have very solid evidence for tyrannosaurs getting to adult size in two decades and big hadrosaurs growing up in half that time or less. Inertial homeothermy is an insufficient explanation because these critters were growing quickly even when they were too small for inertial homeothermy to have done them any good.
We don't know if erect stance is a red herring or not; the causal connection with endothermy is not completely clear, but the circumstantial evidence is pretty good. We don't know whether ancestral crocodylomorphs and rauisuchians were endothermic or ecothermic; if it is naive to assume blanket endothermy for every taxon with erect limbs, it is equally naive to assume blanket ectothermy for every extinct archosaur until proven otherwise. We do know that at least some of those erect-limbed archosaurs grew relatively quickly, and that many ancestral crocodylomorphs were long-legged terrestrial cursors.
The calculations about sauropods being ectotherms because of food intake limitations were flawed in two ways. First, as David Marjanovic explained, they assumed ridiculous handling times for the food. Sauropods not only didn't chew, they couldn't chew. Second, that paper (Weaver 1989) said explicitly that Brachiosaurus had a skull the size of a giraffe's. As Greg Paul has pointed out, this is doubly flawed--not only is a Brachiosaurus skull much, MUCH bigger than a giraffe skull, but it is basically all mouth, whereas giraffes use a crappy little cropping unit down at the front of the snout (and chew their food TWICE, being ruminants).
I suspect that the pro-endothermy talk has quieted down a bit lately because the focus of the science has shifted. Endothermy is slippery to begin with (in terms of how you define it and the number of ways there are to be endothermic) and hard to test. Growth rates, nesting patterns, intra- and interspecific interactions are all directly testable, even if some of them require waiting for the taphonomic lottery to come in.
I'm liking the long sacs on the wing leading edges. They seem necessary, for aerodynamic reasons, in a big flyer. We can imagine them deflating when the wing is folded, and the animal gulping air to inflate them before taking off.
I have argued this point before. Tidal lungs are ancestral. Flow-through lungs cannot possibly work without airsacs. Therefore there had to have been animals with both airsacs and tidal lungs. What is the flaw in this reasoning?
Endothermy is not neccesary for any of these. The highest growth rates known are in altricial birds that are ectothermic at the time.
Please. That conclusion goes way, way beyond any data.
They've still got a narrow wing patagium instead of a continuous surface from ankle to finger-tip, I note. Wonderful paper, though, and great coverage, Darren.
You know, pterosaurs with wingsacs would probably have wings that looks a lot more like an airplane's wings. Rather odd.
Very interesting. Its interesting how pterosaurs combine avian and crocodilian features in their breathing system; air sacs and pectoral musculature associated with the respiratory system.
I have a question though. The picture after "Pterosaurs get SASSy" shows an asymmetrical distribution of air sacs on the wing; I'm assuming one is for ornitocheiroids and the other is for azhdarchoids?
Tidal lungs are ancestral. Flow-through lungs cannot possibly work without airsacs. Therefore there had to have been animals with both airsacs and tidal lungs. What is the flaw in this reasoning?
The "flow-through" part means extrabronchial airflow, which means that the chambers of the respiratory system have to have connections to each other instead of just being connected by the bronchi (the way our lung alveoli are). It is often overlooked that in crocs the septa between adjacent lung chambers are often perforate, which allows at least the potential for extra-bronchial airflow. By phylogenetic bracketing, the null hypothesis is that such potential is primitive for archosaurs.
Also, at some point it becomes counterproductive to talk about lungs vs. air sacs, as if air sacs are something completely different. Many sauropsids have heterogeneous lungs, with some regions of dense parenchyma (more like what we think of as lungs) and some regions of sparse or no parenchyma (what we think of as air sacs). Birds just take this heterogeneity to an extreme. The lungs of crocs are not homologous with only the lungs of birds; they're homologous with the entire lung/air sac system of birds.
Pretty much all of that is in Steve Perry's papers going back to the 1980s; I bring that up not to make you feel bad but just to point out that credit lies elsewhere.
So to answer your question, it did not have to be tidal lungs without air sacs -> tidal lungs with air sacs -> flow-through lungs with air sacs. The lung heterogeneity (differentiation of parts of the lung into what we call air sacs) and flow-through breathing (extra-bronchial airflow) could have both increased together incrementally. Such an evolutionary transition is not easy to break down into neat bins of no air sacs/air sacs and tidal/flow-through; at some point the ancestral critter had "sorta" air sacs and "some" flow-through ventilation.
Although the point was perhaps not made clearly in the original post, the reason we think pterosaurs had a substantially bird-like respiratory system is not just that they had pneumaticity, but that the patterns of pneumaticity we see in them are diagnostic for many of the same air sacs as we find in birds (ditto for sauropods and theropods). So we don't think that the incremental transition from a more croc-like respiratory system to a more bird-like respiratory system happened within any of those clades; we (or at least I) think it was mostly complete before they diverged. Admittedly that leaves the problem of when the air sac system first evolved, and what kinds of evidence we might use to document the transition. All I can say there is that it is a genuinely hard problem, but that several different groups are working on it.
I knew that if I made some bold statements about endothermy I would likely generate some interesting responses. Thanks to all for insightful and interesting comments. I only intend to respond briefly, but here goes...
-- Jerry Harris notes that 'differential inflation of certain regions of the SASS' doesn't seem permitted, given what we know of the air-sac system. He's right â I'll have to query the authors on this point.
-- Paul Barrett argues that the term 'endothermy' is specially associated with 'a specific set of metabolic processes', the existence of which can never be demonstrated for fossil taxa. This is true but, even so, the term endothermy is universally applied to those organisms capable of generating and maintaining heat internally. Ergo, if it seems reasonable to conclude that extinct archosaurs did generate and maintain heat internally, use of the term endothermy would still be correct.
-- Jura notes that some of the evidence noted as supporting endothermy is problematical. As Matt Wedel has already said above, sustained rapid growth in members of all dinosaur lineages is now well supported (e.g., Erickson et al. 2001, 2008, De Ricqlès et al. 2003, 2008, Cooper et al. 2008, Klein & Sander 2008, Lee & Werning 2008, Sander & Clauss 2008) and this evidence better fits an endothermic model than an ectothermic one. Erect stance is not 'a red herring': it's circumstantial evidence, and all animals that have erect gaits have been shown to exhibit sustained elevated growth rates. It's no use using erect-limbed sphenosuchians and rauisuchians to 'demonstrate' that the erect limbs of these taxa correlate with ectothermy, as the metabolic status of these animals is uncertain and in fact histological data suggests that they had elevated metabolic rates (De Ricqlès et al. 2003, 2008). Pareiasaurs are not erect-limbed.
In response to Jura's suggestion that it's better to 'look at this data as equivocal, since that's what scrutiny of the data has shown it to repeatedly be', I would agree that we should be conservative where appropriate (and note that my statements are couched in at least some cautionary language), but otherwise I don't understand you can say this, given what's been published.
And, on the subject of cautionary language, Sven's comment that inferring endothermy is a conclusion that 'goes way, way beyond any data'... err, no it isn't! All data indicates that the maintenance of erect gait, erect neck carriage and elevated growth rate are indeed correlated with endothermy.
Finally - good to see that the subject of ornithodiran endothermy still makes people angry and vocal. Did I also mention that I'm religious?
Just kidding.
Refs - -
Cooper, L. N, Lee, A. H., Taper, M. L. & Horner, J. R. 2008. Relative growth rates of predator and prey dinosaurs reflect effects of predation. Proceedings of the Royal Society B 275, 2609-2615.
De Ricqlès, A., Padian, P., Knoll, F. & Horner, J. R. 2008. On the origin of high growth rates in archosaurs and their ancient relatives: complementary histological studies on Triassic archosauriformes and the problem of a "phylogenetic signal" in bone histology. Annales de Paléontologie 94, 57-76.
- ., Padian, K. & Horner, J. R. 2003. On the bone histology of some Triassic pseudosuchian archosaurs and related taxa. Annales de Paléontologie 89, 67-101
- ., Padian, K., Horner, J. R., Lamm, E.-T. & Myhrvold, N. 2003. Osteohistology of Confuciusornis sanctus (Theropoda: Aves). Journal of Vertebrate Paleontology 23, 373-386.
Erickson, G. M., Curry Rogers, K. & Yerby, S. A. 2001. Dinosaurian growth patterns and rapid avian growth rates. Nature 412, 429-433.
- ., Curry Rogers, K. & Yerby, S. A. 2001. Dinosaurian growth patterns and rapid avian growth rates. Nature 412, 429-433.
Klein, N. & Sander, M. 2008. Ontogenetic stages in the long bone histology of sauropod dinosaurs. Paleobiology 34, 247-263.
Lee, A. H. & Werning, S. 2008. Sexual maturity in growing dinosaurs does not fit reptilian growth models. Proceedings of the National Academy of Sciences 105, 582-587.
Sander, P. M. & Clauss, M. 2008. Sauropod gigantism. Science 322, 200-201.
Just what is it that is supposed to make insisting on ectothermy a conservative position? To be scrupulously conservative would be to have no position on the subject. If you say there's not enough evidence, then an assertion of ectothermy amounts to speculation.
If we're just a little pedantic, it refers only to the way in which the temperature is generated.
Which means that (crown-group) birds and mammals are not capable of having metabolic rates that shift with ontogeny, as occasionally proposed for big dinosaurs in general and sauropods in particular.
Indeed. Now, however, consider the body shape of a sauropod: long neck, long tail, long limbs (relatively longer than an elephant's, though AFAIK the deinotheres got there), and then the air-sac system. How does such an animal conserve heat?
And again, the argument by Paul & Leahy is relevant: such a large animal cannot avoid generating lots of heat. The two papers even quantify that.
Science can't prove anything anyway, it can only disprove -- and it can use the principle of parsimony.
---------------------------------------------
No, speculations. A theory is something much bigger.
<tip, tip, tip>
That's twenty meters. Wow.
---------------------------------------------
Eh... and what about Deinosuchus needing 50 years?
I have a lot of things to say about that book chapter. I wanted to write a lengthy post about it to the Dinosaur Mailing List. I suppose a couple weekends from now I'll get around to it. <sigh>
I'll happily grant, however, that Apatosaurus growing to adult size in 10 years most likely is an overestimate (of the speed, respectively underestimate of the time).
Like what? Dinosaurs remain the only sauropsids in the Late Cretaceous North Slope of Alaska as well as in Kakanaut in Northeast Siberia (I heard a talk by Pascal Godefroit about the latter last week).
---------------------------------------------
To be fair, I don't think this takes longer than doing all of the chewing at once...
---------------------------------------------
That both air-sac systems and flow-through lungs must have evolved gradually.
Intermediate lungs (with a respiratory and a ventilatory part) occur in many extant nonavian sauropsids, as explained above, and don't crocodiles have cross-current exchange, which is already a step closer to the bird condition than the mammalian condition is? Anyway, there's a nice chapter on all this in the Ostrom Symposium volume, which should be available for a (comparatively...) cheap price now.
Ectothermic, yes -- but tachymetabolic. (And unable to switch that off, because it's a property of the cell membranes, as mentioned.)
I'd say it's the most parsimonious option.
---------------------------------------------
I was told by someone who works on Plateosaurus biomechanics that P. has shallow pneumatic fossae on its vertebrae -- just no foramina.
---------------------------------------------
:-D :-D :-D
Frankly, I think it still comes from 1) insisting that dinosaurs should be called "reptiles" and 2) then going on to treat the name "reptile" in a typological way. In reality, phylogenetic bracketing supports ectothermy and bradymetaboly for the first crown-group archosaur, but is ambiguous on everything between that and the first crown-group bird; all else being equal, this makes all hypotheses that fit this picture -- the first ornithodiran being fully endothermic, homeothermic, tachymetabolic and tachyaerobic; all four of these traits evolving in the origin of crown-group birds; the transition being so gradual that it takes up the whole available space; the several traits evolving one after the other instead of all at once; and everything in between -- equally parsimonious. This means there is no default state that one could assume on the question of what thermoregulation pterosaurs or sauropods had before one looks at or in the bones.
I'd like to expand on David's point about Alaska's dinosaurs, because it's not often I'm able to do so. :-)
So far, there are something like ten dinosaurs known from the Colville River (all of them also found in Alberta). There are no crocodilians known, and there are no lizards or amphibians, either. The climate would have been about what it is in modern-day Washington state. Certainly not freezing, but not tropical, either. Bizarrely, Alaska was actually farther north than it is today.
There's a question as to whether our dinosaurs migrated from Alaska to Alberta, akin perhaps to caribou or reindeer. It's certainly not out of the question, but I don't really buy it. Troodon is the most common theropod up there, outnumbering other theropods by something like 2:1, and it has enormous eyes (like Laellynasaura) that suggest it was pre-adapted to the darker conditions in Alaska. Last time I checked, we have the following genera up there:
Albertosaurus, Dromaeosaurus, Troodon, Sauronitholestes, Edmontosaurus, Edmontonia, Pachyrhinosaurus, ?Anchiceratops, Alaskacephale, a lambeosaurine hadrosaur, and an ornithomimosaur.
All of the theropods are known from teeth. Alaskacephale is known from a partial skull and is, so far, the only uniquely Alaskan genus, although its fragmentary nature raises questions about its validity. Sullivan named it in his recent (2006) review of the Pachycephalosauridae.
"Frankly, I think it still comes from 1) insisting that dinosaurs should be called "reptiles" and 2) then going on to treat the name "reptile" in a typological way."
That's why I call crocodilians, birds, non-avian dinosaurs, pterosaurs, and crurotarsians archosaurs rather than reptiles, and put them in a group on par with mammals. Even from a cladistic standpoint (yes, I do use cladistics), one could just call archosaurs a crown group.
Perhaps ornithodirans and crurotarsians both had the advanced traits of endothermy. We know now that all ornithodirans were warm-blooded, and it appears most crurotarsians would be as well, with crocodilians becoming secondarily ectothermic to become more successful in their aquatic niches. It would also explain why the crurotarsian and ornithodiran archosaurs dominated the Mesozoic and shoved most of the basal archosaurs aside, because they were warm-blooded (and for those of you who think crocs can't compete with dinosaurs...well, sebecosuchians and other terrestrial species disagree with you).
"
That's twenty meters. Wow."
No clue what you just said there.
"But once again, these are just theories."
Sorry then, hypothesis. Besides, you try going to a natural history museum and asking them to cut up sauropod bones.
As for all the talk of "x does not indicate endothermy" or "y does not indicate endothermy", the only way we can definitively prove endothermy is if we go back to the Mesozoic and stick a thermometer in the animal's mouth. However, we can be very sure about the possibility of endothermy in animals if they have several traits, such as...
A. Erect gait. Every endothermic animal alive today pretty much has erect gait (not counting gigantotherms and those who use rete mirabilia)
B. Hair or archosaur fuzz. Mammals have hair, and are warm-blooded, birds have feathers, and are warm blooded, and extinct groups that we would expect to be warm blooded, such as pterosaurs and non-avian dinosaurs, have archosaur fuzz as well.
C. Four-chambered heart. Rather difficult to observe if not preserved via a conservation lagerstatten. But so far, four-chambered hearts seem to be indicative of endothermy.
D. Flight. You need to be rather hyper in evolutionary terms in order to fly. Even insects that take flight are rather "hot-blooded" compared to their more terrestrial kin. And so far every tetrapod group that took to the skies has been warm-blooded.
"All of the theropods are known from teeth. Alaskacephale is known from a partial skull and is, so far, the only uniquely Alaskan genus, although its fragmentary nature raises questions about its validity. Sullivan named it in his recent (2006) review of the Pachycephalosauridae."
Once again, the Pachycephalosauridae frustrate us with their weirdness. In addition to us still confused over what exactly did they do with their skulls, pachies also seemed to be one of the only groups diversifying at the end of the Cretaceous. Even if you agree with Horner that Dracorex, Stygmoloch and Pachycephalosaurus were the same species, you still have Stegoceras and Sphaerotholus...in North America alone. Some suggest that pachycephalosaurs were the dinosaurian equivalent of Caprinae, being made up of a variety of mountain-loving and polar species. Just a thought, but perhaps the reason they were diversifying in the Maastrictian was due to them being better adapted to thinner atmosphere as the world was being altered by the Deccan Traps. Just a thought.
"This might sound bizarre but it's present in many large modern birds, including vultures, pelicans and bustards."
And screamers, don't forget screamers. Hence the reason why the people of the region avoid them...so does that mean we could have colossal flying screamer in the future? Yeesh.
"It's no use using erect-limbed sphenosuchians and rauisuchians to 'demonstrate' that the erect limbs of these taxa correlate with ectothermy, as the metabolic status of these animals is uncertain and in fact histological data suggests that they had elevated metabolic rates"
Agreed. In fact the evidence looks like its pointing towards endothermy in crurotarsians, and then only secondarily developing cold-bloodedness. And of course there's the fact that crocs have a four-chambered heart in the embryonic stage, and later lose it.
"And, on the subject of cautionary language, Sven's comment that inferring endothermy is a conclusion that 'goes way, way beyond any data'... err, no it isn't! All data indicates that the maintenance of erect gait, erect neck carriage and elevated growth rate are indeed correlated with endothermy.
Finally - good to see that the subject of ornithodiran endothermy still makes people angry and vocal. Did I also mention that I'm religious?"
Aw dang it! Now I have to edit my list...
E. Elevated growth rate
F. Erect neck carriage
Anyway, provoking discussion is always good.
"or a system of balloons,"
Sounds like this one might be pretty much spot on, though... not that systems of balloons are a novelty specific to Mesozoic dinosaurs and pterosaurs.
This is much more reasonable than Professor Waxman's intended method...
It's unfortunate that Unwin does not discuss what Jim Cunningham found to be constraints on pterosaur size. We do know that it was based off of Quetzalcoatlus and that Unwin at least preferred weights closer to 50 kg than 250 kg. Who knows if the 15-20 m estimate will change with a substantially heavier mass, but surely Quetzalcoatlus and Hatzegopteryx were not flying on the edge of a theoretical limit.
Stephen Baxter, who has a doctorate degree in aeroengineering, included a 100 meter pterosaur in his book "Evolution" - presumably as the result of artistic license.
Re. ectothermy in dinosaurs being the 'conservative' view, this can be seen in a lot of 'popular' works right through the 1980s (which is not so long ago for some of us). F'rinstance, David Norman's stuff was very popular with primary school kids (and their teachers, e.g. my sister) in the UK and Australia, and thus quite influential in forming 'default' ideas of what dinosaurs were like. And meanwhile there's Bakker posing as a heretic, differing from 'orthodoxy' on a lot of technicalities but 'hot-bloodedness' was the biggy, at least in the popular imagination.
Sea snakes are one example of a group with a terminal saccular portion of the lung. From this very simple geometry, how hard would it be for the alveolar portion to evolve a longitudinal septum and valves, creating a one-way flow-through lung? (In the real world, they do a lot of gas exchange through the skin so lung efficiency is not really limiting)
And in contrast to Alaska, Victoria's polar dinosaurs were accompanied by temnospondyls and some kind of lepidosaur. Maybe not quite so cold down under? (But temno's were long-extinct up north and the lepidosaur may have been a cold-adapted sphenodontid, so maybe it's not a fair comparison)
Sea snakes are one example of a group with a terminal saccular portion of the lung. From this very simple geometry, how hard would it be for the alveolar portion to evolve a longitudinal septum and valves, creating a one-way flow-through lung?
Quite a few snakes have this (maybe all?). Brattstrom had a paper in the 50s on "The air sac in snakes". The reigning hypothesis is that the very long terminal saccular portion serves as an air reserve to keep the animal going while the pharynx is blocked by something immense that it is trying to swallow.
As far as evolving that into a one-way flow-through system.... I think the make-or-break would be evolving perforated septa between the chambers in the anterior parenchymal part of the lung. If they did that, then contracting the posterior sac portion at the end might squirt some air from chamber to chamber through those septa. Presumably the chambers so ventilated would experience elevated gas exchange relative to those relying on simple diffusion, and that would be the a step down the path to bird lungs in snakes. I don't think valves are necessary or likely; birds do all their sophisticated air-streaming without any physical valving at all.
How likely is it? Snakes have had quite a while to evolve the system and haven't, so I suppose that either there is some kind of developmental constraint or those oddities that have turned up with perforated septa (it has to happen at some low frequency) haven't been able to glean any selective advantage from it. Still, it's an interesting thought experiment.
One thing that always bugged me about the Ruben et al. contention that theropods had diaphragms and couldn't have evolved air sacs without getting hernias--besides the appalling neglect for taphonomy, anatomical consistency, evidence that they already had air sacs, etc.--was the unstated dismissal of reversal. Surely a hypothetical diagphragmatic theropod could revert to the ancestral diaphragm-less condition and then evolve in a new direction.
Since I'm hogging the comments anyway, it's past time I said thanks, Darren, for the shout-out. Nice post as usual. Keep 'em coming!
Well, here's hoping I don't break the comments.
Darren wrote:
Does it? How many actual ectotherm growth rate studies have their been? How many that focused on actual reptiles? Most of the data comes from Case's 1978 review of growth in vertebrates, wherein Case mentions: "Reptilian growth is difficult to standardize because of its extreme individual and temporal variations, as the growth often falls to zero during unfavorable periods.
...I have calculated it only during the period wherein the absolute rate of growth is relatively constant. For reptiles, this period was sometimes not obvious."
Despite being well cited, the reptile data was not the most rigorously obtained.
I find it interesting that this is the first group of non-dinosaurian reptiles to actually be considered in the pro-endotherm camp (I suppose the second would be plesiosaurs). I find it more than a little convenient that these guys are now considered to be possible automatic endotherms as well, just because they share all the same characteristics seen in dinosaurs, which are presumed to lead to automatic endothermy; yet despite the fact that their descendants are bradymetabolic.
Oh, but there's a way around that right? Crocodylians are just secondarily bradymetabolic, like naked-mole rats. Seems nice and simple until one peruses the taxa in question. I mean, when did this alleged retroversion occur? After the split from other pseudosuchians? (so Protosuchus, and Sphenosuchus are bradymetabolic then?), at the Pholidosaur split (so sebecosuchians and Uberabasuchus are bradymetabolic?) Perhaps at the base of Eusuchia (Quinkana and Pristichampsids are now the only super-active bradymetabolic taxa)? This argument really doesn't seem that parsimonious.
Also, for the record, Pareiasaurs were erect limbed (see Romer, 1956, for a general, though hardly extensive, overview), and extant crocodylians (depending on species) are also erect limbed (hind limbs, at least).
Wait, when did erect neck carriage come into play. Should I start to refer to the little dwarf caimans of South America as automatic endotherms now? I mean Paleosuchus palpebrosus is renowned for carrying its "dog-like" head high over the shoulders. All these criteria just seem like arbitrary associations for automatic endothermy. I never see any real information to explain why this must be so.
Dr. Jack wrote:
If only that were the way. Whenever the pro-endotherm stance is taken, it is never in regard to myogenically produced, functional endothermy like that seen in lamnids, varanids Dermochelys, and insects. It's always in regards to the metabolically wasteful, viscerally produced, "can't shut off" automatic endothermy that is seen in birds and mammals.
David M. wrote:
There are issues with how that paper handled the Deinosuchus growth curve. Some of it was mentioned onlist by HP Adam Britton, and others were mentioned in David Schwimmer's (2002)Deinosuchus book (i.e. the existence, and fidelity of LAGs in what would have been an equatable climate, and whether, or not these were even annual). As it stands, the growth curve is actually too slow for extant crocs.
If anything, it is the pro-endotherm camp that is suffering from this mentality. Mainly the view that "reptiles" are incapable of sustained activity, parental care, high growth rates, polar existence, etc. Dinosaurs "had" to be automatic endotherms in order to deal with this perceived problem. Find a "classic" reptile in an area like this (such as crocodylians, and chelonians living both in the Northern Polar regions [Axel- Heidelberg island], and Southern polar regions [Victoria]), and it's likely ignored, rationalized (it was "unseasonably" warm at the time), or in those very rare instances, co opted into a pro-endotherm view (i.e. the recent trend in pseudosuchian physiology).
This is why I love reading this blog. Reality, including realities that were, is infinitely more interesting than fantasy.
The polarmost modern squamates and lissamphibians are in northern Scandinavia - surely that's colder than Cretaceous Alaska?
For the record, I am not and was not arguing that ornithischians or sauropods were ectothermic. I really don't know, and strongly suspect these creatures had metabolisms rather different from anything alive today. I am stating that the evidence for ornithischians being endothermic is at best ambiguous, with most of the facts being cited as supporting evidence being either trivial or also true of some ectotherms. For sauropods also the available evidence is way too sketchy for any conclusions to be drawn.
My earlier reference to the calculation of whether or not sauropods could consume sufficient calories to maintain a truly endothermic metabolism were not intended to suggest that I belive this to be a killer blow to sauropod endothermy - only that there are significant challenges. The comment about their not chewing is relevant, the one about the head size I think less so, since I saw the calculation done in a manner that negated that. Please also note that there was a wide margin for error, so the calculations don't just have to be wrong, they have to be very very wrong for the larger sauropods to have been able to consume sufficient calories per day. Maybe the calculations are very wrong, but as it stands I still think they present a challenge to the idea that sauropods were truly endothermic that has not been fully dealt with.
As for using phylogenetic bracketing - that isn't evidence, it's what is resorted to in the absence of evidence.
So far I think that the evidence for endthermy in pterosaurs and some theropods is fairly convincing. The evidence for it in other archosaurs is moot; even for ornithischians and sauropods it is either ambiguous or having approximately equal counter arguments, and for the other groups it is mostly poor.
Brief response to Jura: I think it's all too easy for those dismissing (yes, dismissing) the possibility of endothermy in Mesozoic archosaurs to say that a 'pro-endothermy' stance reflects a typological view: a typological view in which non-ornithodiran reptiles (like extant crocs and squamates) don't practise complex behaviours, are capable of limited exercise etc. There is no indication that this is true, at least not among the modern proponents of endothermy, and it's a straw-man argument. Padian & Horner (2004) presented a lengthy discussion of how views on metabolism have been affected by typology: it's clear that the 'pro-ectothermy' position is the biased one.
On the subject of evidence for endothermy in non-ornithodiran archosaurs, your implication that this is invoked because it's 'convenient' is naïve: it's a conclusion based on published data, not on intuition. Similarly, the possibility of secondarily-acquired bradymetabolism in crocodylians is not speculative intuitive guesswork as you imply, but based on histological data and developmental biology. I base these contentions on De Ricqlès et al. (2003, 2008) and Seymour et al. (2004). If you haven't read these papers you should do so.
Pareiasaurs have a semi-erect hindlimb gait (like rhynchosaurs and other archosauromorphs) but a sprawling forelimb; they were not fully erect-limbed (I've somehow lost my folder of pareiasaur literature so cannot cherry-pick comments right now). The presence of a constantly maintained fully erect limb posture (as seen in dinosaurs and extant endotherms) is not conclusive evidence for endothermy, but suggestive. Similarly...
Erect neck carriage: ancestrally, all crown-group tetrapods (including crocodylians) carry the neck extended relative to the dorsal column. But the angle is shallow, and constantly maintained fully erect neck carriage (present in dinosaurs as well as extant endotherms) does seem correlated with endothermy.
Let's be clear: the evidence for endothermy in ornithodirans is by no means conclusive or utterly convincing. The point is that what evidence we have, from such things as bone histology, growth rates, limb posture etc., is highly suggestive of endothermy. Highly suggestive to the extent we can say: they probably were endothermic. So far, the argument against this amounts to: "but they might not have been, and the evidence is not conclusive". Well, duh.
Refs - -
De Ricqlès, A., Padian, P., Knoll, F. & Horner, J. R. 2008. On the origin of high growth rates in archosaurs and their ancient relatives: complementary histological studies on Triassic archosauriformes and the problem of a "phylogenetic signal" in bone histology. Annales de Paléontologie 94, 57-76.
- ., Padian, K. & Horner, J. R. 2003. On the bone histology of some Triassic pseudosuchian archosaurs and related taxa. Annales de Paléontologie 89, 67-101.
Padian, K. & Horner, J. R. 2004. Dinosaur physiology. In Weishampel, D. B., Dodson, P. & Osmólska, H. (eds) The Dinosauria, Second Edition. University of California Press (Berkeley), pp. 660-671.
Seymour, R. S., Bennett-Stamper, C. L., Johnston, S. D., Carrier, D. R. Grigg, G. C. 2004. Evidence for endothermic ancestors of crocodiles at the stem of archosaur evolution. Physiological and Biochemical Zoology 77, 1051â1067.
Darren, this is a minor point in this endothermy discourse, but I was wondering about what you wrote concerning neck posture:
and
If we ignore birds, which Recent mammalian endotherms (apart from us humans) carry their necks 'constantly fully erect'? Off the top of my head, it would seem to me that most mammals do, in fact, carry their necks at relatively shallow angles for most of the time. Is there some generally agreed-upon point after which carrying the neck at a 'shallow angle' becomes carrying the neck 'fully erect' (functionally speaking)? Could you please give me the reference(s) for those data on the correlation between erect neck carriage and endothermy?
"Oh, but there's a way around that right? Crocodylians are just secondarily bradymetabolic, like naked-mole rats. Seems nice and simple until one peruses the taxa in question. I mean, when did this alleged retroversion occur? After the split from other pseudosuchians? (so Protosuchus, and Sphenosuchus are bradymetabolic then?), at the Pholidosaur split (so sebecosuchians and Uberabasuchus are bradymetabolic?) Perhaps at the base of Eusuchia (Quinkana and Pristichampsids are now the only super-active bradymetabolic taxa)? This argument really doesn't seem that parsimonious."
It seems that crocodilians experimented with erect gaits and endothermy several times throughout their evolutionary history, such as the sebecosuchians, the mekosuchians, and the pristichampsids. However, none of these lineages have survived to the present day, and thus only the more conservative, ectothermic crocs managed to make it into the present. Wow...I guess the meek really do inherit the earth.
"Let's be clear: the evidence for endothermy in ornithodirans is by no means conclusive or utterly convincing. The point is that what evidence he have, from such things as bone histology, growth rates, limb posture etc., is highly suggestive of endothermy. Highly suggestive to the extent we can say: they probably were endothermic. So far, the argument against this amounts to: "but they might not have been, and the evidence is not conclusive". Well, duh."
Agreed. It appears most likely that all ornithodirans were endothermic, there is no way of actually completely proving it other than than getting a time machine, going back to the Mesozoic, and sticking a thermometer in their mouth...or their other entrance.
"On the subject of evidence for endothermy in non-ornithodiran archosaurs, your implication that this is invoked because it's 'convenient' is naïve: it's a conclusion based on published data, not on intuition. Similarly, the possibility of secondarily-acquired bradymetabolism in crocodylians is not speculative intuitive guesswork as you imply, but based on histological data and developmental biology. I base these contentions on De Ricqlès et al. (2003, 2008) and Seymour et al. (2004). If you haven't read these papers you should do so."
Yes, agreed with that as well. There are numerous examples of active, fast-moving, erect-limbed crurotarsians in the fossil record, whether it be the terrestrial crocodilians, or the rauisuchians and kin of the Triassic.
"Mainly the view that "reptiles" are incapable of sustained activity, parental care, high growth rates, polar existence, etc."
Who ever said this? From looking at modern animals, it does appear that endotherms tend to have more complex behavior than ectotherms, but that doesn't mean that ectotherms cannot have complex behavior. Look at crocodilians, and their courtship rituals, how nile crocs will sometimes work together to catch fish, and how they build nests for their young. It does appear, however, that archosaurs have a tendency to have more complex behavior than squamates, but that is only looking at modern species.
But as for polar ectotherms...don't really agree with you there. The only polar amphibian or non-archosaurian reptile that really lives far up north is the Wood Frog, and it survives by being frozen over a lot of the time.
It is also really easy for crocodilians to develop endothermy, especially from an ontological view. Crocodilians start out with a four-chambered heart, one of the hinting signs of endothermy, in the womb, and then lose it and become three chambered. Rather unusual, but then a simple quirk in development could lead to a four-chambered heart, a hyper crocodile, and then endothermy.
"It's unfortunate that Unwin does not discuss what Jim Cunningham found to be constraints on pterosaur size. We do know that it was based off of Quetzalcoatlus and that Unwin at least preferred weights closer to 50 kg than 250 kg. Who knows if the 15-20 m estimate will change with a substantially heavier mass, but surely Quetzalcoatlus and Hatzegopteryx were not flying on the edge of a theoretical limit."
I think that is what he was talking about in the book, I'm not sure, I'll have to pull it out.
The lizard [i]Zootoca/Lacerta vivipara[/i] reaches similar latitudes in Scandinavia. (Now, certainly, Scandinavia is unreasonably warm for its latitude by modern standards, but modern standards are very cold by Mesozoic ones.)
Dartian asks...
I'll have to be cryptic here and say 'wait for the paper', sorry. Seriously, an ms on this topic is due to appear soon. It WILL be discussed here at Tet Zoo :)
Metalraptor:
Andreas already mentioned the viviparous lizard Zootoca (a.k.a. Lacerta) vivipara. But there are other 'herps' that are found at, or even north of, the Arctic Circle in Scandinavia: common frog Rana temporaria, moor frog Rana arvalis, common toad Bufo bufo, and adder Vipera berus.
Darren:
Quantitative data are forthcoming? Yay!
Just considering some of those bits of evidence again:
Erect gait - most extant endotherms have an erect gait, but there are exceptions, also most extant ectotherms do not have an erect gait, but again there are exceptions. While there does seem to be a general corelation among extant tetrapods, there doesn't seem to be any compelling reason why erect gait should be tied to endothermy, and the exceptions demonstrate it isn't. Erect gait would be a much more convincing indicator of endothermy if there was a convincing biological connection between the two.
Erect neck carriage - again among extant tetrapods usually but not always a feature of endotherms. In deed to me this is one of the poorest pieces of evidence put forward. Darren you state that a "constantly maintained fully erect neck carriage (present in dinosaurs as well as extant endotherms) does seem correlated with endothermy." But why? What biological reason is there? It would be way more convincing if we could say erect neck carriage is usually associated with endothermy because... The fact that object A posses properties 1 and 2, is not evidence that object B because it also posses property 1 must also posses property 2, unless you can demonstrate some connection between properties 1 and 2 that means they tend not to be independent of each other. This is simple logic.
Growth rate - is potentially more telling, and I used to find it rather convincing, but once again there are exceptions. Growth rate and continuity tells us something about the environment in which bone was laid down, but whether that environment was primarily maintained by the animal's metabolism or by the external environment is not revealed. In our time there is a good (but not complete)correlation between rapid and fairly constant growth rates and endothermy, whether that applied in the rather different and much warmer environment of the mesozoic is a whole different issue. Seasonal and slow bone deposition would seem to be a better indicator of ectothermy than non-seasonal and fast bone deposition is of endothermy.
Hair, feathers or archosaur fuzz - the use of some kind of insulator does seem to be suggestive of retaining a higher internal temperature than external. While not conclusive, I do find this much more persuasive, since there does seem to be a biological connection, not just a correlation.
Flight - the energetic requirements of flight do tend to support the idea that endothermy could well be connected.
Nasal turbinates - the absence of nasal turbinates has been used as an argument against endothermy in dinosaurs, since it is a feature of most extant tetrapod endotherms, and is known to play a functional role related to endothermy. But the feature is not universal. I would suggest that this is a piece of evidence against endothermy on a par with erect gait as evidence for - i.e. it is at best suggestive. To me the absence of nasal turbinates in dinosaurs etc is not proof of ectothermy, but their presence if discovered would be highly suggestive of endothermy.
Darren you state:
"The point is that what evidence he have, from such things as bone histology, growth rates, limb posture etc., is highly suggestive of endothermy. Highly suggestive to the extent we can say: they probably were endothermic. So far, the argument against this amounts to: "but they might not have been, and the evidence is not conclusive". Well, duh."
I don't think that is very fair. While for pterosaurs and some theropods I would agree that the evidence is "highly suggestive to the extent we can say: they probably were endothermic" - the whole point is that for many of us the evidence does not look anything like that good for the other archosaurs. You imply that some of us are using "but they might not have been, and the evidence is not conclusive" as an argument for ectothermy. That is not so. They may have been endothermic, they may have been ectothermic, they may have run on nuclear power cells (okay, maybe not the nuclear power cells), it just seems that conclusions are being drawn way beyond the evidence - great fun as speculation, not so good as science.
Phylogenetic bracketing combined with special pleading to explain the ectothermy of all extant crocodylians doesn't impress me very much. I have read (and enjoyed) Seymour et al. (2004) and one of the De Ricqlès papers (can't recall which) but I clearly wasn't as wowed by the evidence for endothermy in non-ornithodiran archosaurs or of reversion in crocodylians as you were.
I hope we will one day know what the metabolism of ornithischians and sauropods was. And personally I hope they were endothermic - I would like Triceratops to have been turbocharged - but as of now I think the evidence is just not so strong that "we can say: they probably were endothermic".
Sorry for prattling, and I am consious that all this endothermy/ectothermy argument is distracting from the really good stuff you had to say about pterosaur airsacs. So having had more than my fair share of space (I have the day off work and am desperately trying to avoid doing work in my garden), Im going to shut up now.
Well, now I understand why John Hawks doesn't allow comments on his blog - there simply is not enough time in the day to respond to everything you want to. Thanks for the long comment Mark.
Mark, first off, the presence of one of those traits does not mean that the creature was endothermic, you are right there. But many of these creatures have a combination of said traits, which suggests that they are endothermic. Theropods and pterosaurs have erect gait, erect neck carriage, integumentary structures, flight (in birds and pterosaurs), and a rather rapid growth rate suggest that these animals were endothermic. Ornithiscian dinosaurs have the same except for flight (for integumentary structures, think Psittacosaurus). And while phylogenetic bracketing does not conclusively prove something on its own, the evidence already present plus phylogenetic bracheting suggests that ornithiscians were the same. However, the sheer size of sauropod dinosaurs suggest that they either had a slower metabolism (like living sloths), or they had some adaptation that we do not yet know of in their metabolism. Of course, then again, they used a giraffe skull as a basis for a sauropod skull in those energy experiments as mentioned above, so there may be some problems there.
And thanks for the stuff on other northern herps. But the crocodilians stuff is not pleading. There are features which point to endothermy in crurotarsians, and crocodilians switching between an active and sluggish lifestyle multiple times in their evolution. Perhaps the gap between endothermy and ectothermy is more easily crossed than we thought. However, since the research on Triassic crurotarsians compared to ornithodirans is severely lacking, this matter will have to remain a muddle for another day.
"I don't think that is very fair. While for pterosaurs and some theropods I would agree that the evidence is "highly suggestive to the extent we can say: they probably were endothermic" - the whole point is that for many of us the evidence does not look anything like that good for the other archosaurs. You imply that some of us are using "but they might not have been, and the evidence is not conclusive" as an argument for ectothermy. That is not so. They may have been endothermic, they may have been ectothermic, they may have run on nuclear power cells (okay, maybe not the nuclear power cells), it just seems that conclusions are being drawn way beyond the evidence - great fun as speculation, not so good as science."
Finally, this, I completely agree with this. The evidence does seem to suggest some form of endothermy in all archosaurs, but so far we cannot definitively say one way or the other.
And indeed, there are hadrosaur eggs in Kakanaut.
Leaellynasaura, after Lea Ellyn, the daughter of Thomas Rich and Patricia Vickers-Rich.
I calculated what "66 feet" means, so that I could imagine something. :-)
No, monotremes don't, for example. Endothermy came first.
While insects are never homeothermic*, because almost all are too small for that, they are tachyaerobic (because the tracheae reach every cell directly or almost directly), can therefore be as tachymetabolic as they wish, and are therefore endothermic during flight.
* The nests of social insects as a whole, however, often are homeothermic.
The addition of a little bit of carbon dioxide wouldn't result in a "thinner atmosphere".
They never lose it. They develop a valve (foramen Panizzae) between the⦠um⦠the lung artery and the right aorta or something⦠anyway, it's not in the heart, which is fully four-chambered.
The empirical limit (which has been hit on Pharyngula) is above 2500 comments. Our 50 here don't count.
The absolute height of the head above the shoulders is what counts here. To sustain a head that is, in absolute terms, carried high above the shoulders, you need a strong heart which by itself has a high metabolism.
(This heart also needs to be four-chambered so the lung capillaries don't explode, but crown-group archosaur hearts are already four-chambered by default, judging from phylogenetic bracketing.)
Completely irrelevant. LAGs are endogenic; they occur even in alligators that are kept indoors, and are not unknown in mammals and birds either.
That's a better question!
How does that work?
This was the case in the 1980s and earlier. Nowadays it's just a strawman.
At the PETM, it really was unreasonably warm, warmer than ever since the Cenomanian-Turonian boundary. Maastrichtian Alaska and NE Siberia were considerably colder than early Eocene Axel Heiberg Island. Apparently they were also a bit colder than Aptian-Albian Victoria, but I'm not very well informed about that.
I'm not sure.
Please explain.
I'm sitting in a lab with a huge folder of pareiasaur literature, so if you can tell me which papers to look upâ¦
I don't understand why Jura cited Romer 1956 as if nothing had been done since. There's work by Mike Lee, for instance.
It should be mentioned, though, that most of them -- most clearly Pristichampsus, the biomechanics of which have been studied in some detail -- seem to have been ambush predators, perhaps outright sit-and-wait predators.
Like what?
Why? Why on the planet? Why isn't it the other way around -- the energy required to keep such a body from falling down, let alone moving it, is so high that it can only be provided by a fast metabolism?
Elephants haven't got a slow metabolism.
"It should be mentioned, though, that most of them -- most clearly Pristichampsus, the biomechanics of which have been studied in some detail -- seem to have been ambush predators, perhaps outright sit-and-wait predators."
It should also be mentioned that Pristichampsus is the least derived terrestrial lineage from the crocodilian norm. It is the most like its aquatic ancestors, while other terrestrial croc groups like sebecosuchians were fully terrestrial, and some mekosuchines are thought to even have been semi-scansorial.
"most extant ectotherms do not have an erect gait, but again there are exceptions.
Like what?"
Agreed.
"The absolute height of the head above the shoulders is what counts here. To sustain a head that is, in absolute terms, carried high above the shoulders, you need a strong heart which by itself has a high metabolism.
(This heart also needs to be four-chambered so the lung capillaries don't explode, but crown-group archosaur hearts are already four-chambered by default, judging from phylogenetic bracketing.)"
That's what I've been saying.
"While insects are never homeothermic*, because almost all are too small for that, they are tachyaerobic (because the tracheae reach every cell directly or almost directly), can therefore be as tachymetabolic as they wish, and are therefore endothermic during flight."
That's why I said hotted blooded, rather than endothermic, as they are not true endotherms per se, but their bodies are kept warmer than their flightless kin.
"No, monotremes don't, for example. Endothermy came first."
Even more evidence for the "archosaurs outside of the ornithodirans could have endothermy" argument.
Sorry, I meant I don't understand this... ""
Anyway, it seems like a lot of the "dinosaurs might be ectothermic" stuff that's being said here is due to phylogenetic bracketing, not the other way around. Essentially its saying that "modern squamates are ectothermic, extant crocodilians are ectothermic, and such, so ornithischian dinosaurs had to be ectothermic". Of course you know that lizards are not that closely related to dinosaurs. And there is evidence of at least some form of warm-blooded-ness in crurotarsians. And of course both theropods and pterosaurs are warm blooded, and ornithischian dinosaurs have many similar features to the theropods (I remember one study recently in National Geographic that said that ornithiscians actually grew faster than theropods). The only thing I can think of that ornithiscians are missing are integumentary structures (other than Psittacosaurus and Triceratops). But then again, lack of hair doesn't make rhinos, hippos, and elephants cold-blooded now does it?
Finally, I'm not sure if this has been debunked, but I had thought they had found a "heart" (last I checked people were arguing if it was a heart or a mineralogical concretion) in a dinosaur...specifically Thescelosaurus, an ornithischian.
David wrote:
I wasn't worried about the comment count, so much as the overall comment length. Some of these comments (e.g. my last one) were getting to message board lengths. I'm not sure whether, or not the blogging software was capable of handling long comments, as they are generally rare.
Because it was nearby, and it was late. Most of what I've read from Lee, has had more to do with the chelonian affinities of pareiasaurs (dermal armour, skull design), and less to do with their biomechanics.
True, they don't. They have a metabolism that is only slightly higher than an elephant sized reptile (calculated by Spotila in 1980), which is one line of evidence used to suggest that automatic endothermy, once obtained, is hard to lose.
Again, then Paleosuchus should be considered to have a high metabolism. Or at least a metabolism as high as a similar sized mammal, as that is what carrying it's head that way is often compared too. Paul Willis's work on Quinkana found that it also carried its head high, on its (likely) erect limbs. There are also more than a few elapids that move around with the upper 3rd of their body carried of the ground.
In response to Darren - I've read the papers you've mentioned, and did not come away from them as convinced as you were. I agree with Padian and Horner, that finding one "magical" trait for endothermy, seems pointless, but I disagree that the suite of evidence suggests automatic endothermy in dinosaurs.
The Seymour paper I take particular issue with. All the authors found was evidence that many crocodylomorphs archosaurs were living active lifestyles. They simply stated that activity = automatic endothermy, with no real reason why it should be so, or even if such as stance is valid. I can't help but think that if monitor lizards, and teids (and elapids, racers, other autarchoglossans) were extinct, and all we had to go on was fossil evidence; we would assume automatic endothermy for this group too.
Also they said that they compared croc DNA with fossil croc DNA, which I thought was just weird.
As for the pro-endotherm stance using an outdated view of reptiles being a strawman, I refer you to Bakker's Heresies, Paul's Predatory Dinosaurs of the World, Paul and Leahy's Terramegathermy paper (or one of its derivatives), or really anything regarding metabolism from GSP.
Far from a strawman, it seems to be Paul's key stance on this issue. So much so that he has even attempted re-naming thermophysiological criteria in such as fashion that it would negate any possibility of any reptile ever being considered "warm-blooded" (he basically qualified each term relative to the "standard reptilian condition"). And Paul remains one of the most vocal supporters of "warm-blooded" dinosaurs.
Just because other people do not know the correct definition of endothermy, this does not mean that the majority view that it's essentially the same as 'elevated body temperature', as some commenters have argued, is correct. Precision is immportant people - science is not a democracy - it's about getting the facts straight and the p values right. There's too much arm waving and not enough hypothesis testing in palaeobiology in general. The more quantitative and testable we try to be the better.
Jura said: "How many actual ectotherm growth rate studies have their been? How many that focused on actual reptiles? Most of the data comes from Case's 1978 review of growth in vertebrates..."
This statement is just flat-out wrong. See recent papers by Castanet, de Buffrénil, Laurin, etc. I don't have time to cite all the references, but just do a Google Scholar search for the terms "skeletochronology" and "Sqauamata" (or Amphibia). Also check out these references on crocodylians (there are many others that also publish growth curves):
Games, I. 1990. Growth curves for the Nile crocodile as estimated by skeletochronology. pp. 111â121 in Crocodiles: Proceedings of the 10th Working Group Meeting of the Crocodile Specialist Group, Volume 1. IUCNâThe World Conservation Union, Gland, Switzerland.
Hutton, J. M. 1986. Age determination of living Nile crocodiles from the cortical stratification of bone. Copeia 1986:332â341.
Peabody, F. E. 1961. Annual growth zones in living and fossil vertebrates. Journal of Morphology 108:11â62.
Tucker, A. D. 1997. Validation of skeletochronology to determine age of freshwater crocodiles (Crocodylus johnstoni). Marine and Freshwater Research 48:343â351.
Woodward, A. R. and C. T. Moore. 1992. Alligator age determination. Final Report, Study Number 7563, Bureau of Wildlife Research. Florida Game and Fresh Water Fish Commission, Tallahassee, Florida, 20 pp.
gah. I'm way behind on the paleophysiological literature and have no time to catch up. Let me throw out a few thoughts to fuel the discussion (from which I'm learning stuff).
Precision in terminology is indeed important, and these discussions always get muddled in concept-conflation. Endothermy refers only to the source of body heat. Homeothermy refers only to the temporal stability of body temperature. High resting metabolic rates have been called tachymetabolism. The ability to sustain high levels of aerobically supported exercise has been termed tachyaerobic. "Warm-blooded" means absolutely nothing at all. Going back in time and taking a dinosaur's body temperature would tell you only its body temperature--nothing else.
The proximate and ultimate interrelationships among these concepts (and their proximate and ultimate relationships to morphological traits like 4-chambered hearts and flow-through lungs) are complex and poorly understood, in many cases, even in extant animals. I am most receptive to hypothetical linkages that include some sort of mechanism. The presence of turbinates is an example--I can't think of another reason for having them other than reduction of water- and heat-loss concomitant with high respiratory rates (which imply high oxygen demands and therefore high metabolic rates). David's argument that erect neck carriage in a large animal requires high blood pressure, therefore a large, hard-working heart that would itself increase metabolic rate makes sense. I can see the same argument for erect gait requiring constant isometric muscle tension, again helping to cause a high metabolic rate (and therefore requiring the physiological capacity to support that metabolic rate). Insulation is an excellent indicator of (at least limited) endothermic homeothermy.
But I just do not understand the argument from growth rates. Metabolic rates are effects, not causes--a high metabolic rate is not a potentiating engine that powers other high rates. As has been noted, mammals have high resting metabolic rates because they use a lot of energy at rest (pumping ions back across leaky membranes). That ion pumping is an allocation of energy that decreases the energy available for growth--it in no way automatically increases growth rate. Fast-growing altricial birds are not fast-growing because they are "tachymetabolic;" however high their resting metabolic rate is (I do not know of any data offhand), that metabolic rate is caused by their growth rate, not the other way around. I certainly agree that high and constant body temperatures will (all else equal) increase growth rates, but the wasteful energy-spending of endothermy? Not so much. Am I missing something? Can anyone explain the argument from growth rates in any way that includes a plausible mechanistic link?
Finally, ecothermy is a reasonable (and conservative) starting assumption for dinosaurs not because of bracketing (that makes no sense), but because it is clearly ancestral for tetrapods, amniotes, and sauropsids. Endothermy is a specialization, it's hugely wasteful, and its advantages are not so clear as seems to be assumed by most of us endotherms.
Did anyone else notice that the orientation of the pteroid bone is not consistent with what Unwin et al. have been saying about it recently? What gives?
I just noticed that today, Boesse! Made me chuckle.
Alaska's north slope dinosaurs lived way up HERE, not far from the Arctic Ocean. Now, keep in mind that back in the Late Cretaceous, Alaska was actually farther north than it is today. But ultimately, that's not important. What's important is the temperature at the time. Like I said before, according to the plant fossil record, the climate was similar to modern Washington state. More wet than cold, and it might not have snowed very often, but ice would've been a factor. The real area of concern is the darkness--the sun would have set for likely a whole month up there during the winter, so what did the animals live on during that time? And besides that, during the rest of the winter, the sun wouldn't have come out for very long in the day.
The situation is akin to Australia's Early Cretaceous fauna. And David, I know where Leaellynasaura gets its name--I've met Tom and his wife (nice folks!). It's just a hard name to spell off the top of my head. :-)
"But I just do not understand the argument from growth rates. Metabolic rates are effects, not causes--a high metabolic rate is not a potentiating engine that powers other high rates. As has been noted, mammals have high resting metabolic rates because they use a lot of energy at rest (pumping ions back across leaky membranes). That ion pumping is an allocation of energy that decreases the energy available for growth--it in no way automatically increases growth rate. Fast-growing altricial birds are not fast-growing because they are "tachymetabolic;" however high their resting metabolic rate is (I do not know of any data offhand), that metabolic rate is caused by their growth rate, not the other way around. I certainly agree that high and constant body temperatures will (all else equal) increase growth rates, but the wasteful energy-spending of endothermy? Not so much. Am I missing something? Can anyone explain the argument from growth rates in any way that includes a plausible mechanistic link?"
The correlation between growth rates and size is that endothermic animals have been somewhat noted to grow at a constant rate. Ectothermic animals tend to speed up and slow their grown depending on the available food and temperature.
"Finally, ecothermy is a reasonable (and conservative) starting assumption for dinosaurs not because of bracketing (that makes no sense), but because it is clearly ancestral for tetrapods, amniotes, and sauropsids. Endothermy is a specialization, it's hugely wasteful, and its advantages are not so clear as seems to be assumed by most of us endotherms"
Yes, but realize also that dinosaurs possess many features which are found only in endotherms (erect gait, high neck carriage, etc.) Also note that both theropod dinosaurs and the more distantly related pterosaurs both have endothermy, which in conjunction with the traits noted in ornithischian and other possible dinosaurian ectotherms suggest that they were warm blooded as well. Just because an animal
Essentially if it walks like a chicken, clucks like a chicken, has pennaceous feathers like a chicken, four toes with one of them being reversed like a chicken, and lays eggs like a chicken, its not a lizard, its a chicken (or at the least an avialian archosaur).
Sorry, comment got cut off again. Kept hitting that post button by accident. This is what I meant to say.
Just because an animal doesn't have integumentary structures or is not capable of flight does not mean it is automatically ectothermic.
Endothermy requires both an internal source of heat (whether constantly working muscles, big hard-working viscera, or ion-pumping membranes) AND insulation to hold the heat in (fur, feathers, blubber, or bulk).
Unless I've misunderstood, the argument from growth rates I'm talking about isn't "dinosaurs grew constantly therefore they were endothermic," it's "dinosaurs grew fast, therefore they were endothermic."
Paul Barrett argued "Just because other people do not know the correct definition of endothermy, this does not mean that the majority view that it's essentially the same as 'elevated body temperature', as some commenters have argued, is correct."
Indeed it should be mentioned that ectotherms are capable of maintaining stable elevated body temperatures, maintained by behavioral means. These animals do not possess the evidence for high growth rates, erect gait and sustained muscular activity seen in mammals, birds and dinosaurs though. The evidence indicating stable elevated temperatures combined with other evidence does lead to the conclusion that true endothermy, the internal metabolic production of heat, was the case in dinosaurs.
Again I would say that there is nothinga in our undestanding of endothermy that make it "better" (in the sense of mammal chauvinism) than other metabolic regimes, given that endothermy is widespread in insects and fish.
Unless I've misunderstood, the argument from growth rates I'm talking about isn't "dinosaurs grew constantly therefore they were endothermic," it's "dinosaurs grew fast, therefore they were endothermic."
They grew fast as well, another point for endothermy in dinosaurs.
Can someone point to or summarize such evidence? I have not seen it in this thread.
Gah! That's the assertion I'm not buying!
Love the rant! I have suspected endothermy in all ornithodira for quite some time now. For pterosaurs it's especially clear that they would have needed some kind of thermal control to deal with their wings. Membranes like that can loose or gain energy too fast for comfort.
[problems come from]
So let's revive Dragons! ;-D
(apologies to anyone who sees this as OT)
Dragon is a good word. Let's use it!
I have often (since being a tiny kid able to talk about it) thought that dinosaurs (and others including the extant Komodo already so-named) deserve to be called simply "dragons" as their collective common household name. Many of their scientific names do incorporate 'draco' - or other mythological names such as Quetzalcoatlus.
I suggest the major reason for not (re-)adopting "dragon" historically at the outset was that Owen et al had the idea dragons were mythological only - had never existed - a haughty superior attitude to our ancestors, and wanted the personal kudos of naming all 'fresh, new' stuff (like Marsh and Cope in the later bone wars out west). 'Dinosaur' was thus a neologism that staked the scientific establishment's monopolising claim to a 'new' field. Thus the word 'dinosaur' is itself a fossil, relic of an age of disconnection, imperialism and arrogance.
But as Adrienne Mayor points out in her book The First Fossil Hunters" ancient Greeks, Romans and others had, long before, found and sought to interpret fossils. Many classical monster legends may come from such real finds.
Using "dragon" now would have the advantage of humbly and humanely reestablishing cultural continuity with most of humanity who AFAIK throughout history and worldwide generally have retained a concept of "dragon", whether from spectacular unfamiliar bones they found personally in their own generation or traditions from those who had earlier known such things. It would build a bridge linking science to the arts and humanities. Accounts of local 'dragons' in classical legend and folklore, if not simply dismissed as fantasy, might usefully point to fossiliferous areas previously known and worth re-examining.
(We could still distinguish between 'real dragons' and 'made-up dragons'. But I suspect that distinction would have to remain fuzzy, since all dinosaurs and extinct creatures depend on human imagination to give them form. The distinction is largely a matter of the degree to which any discoverer's/reconstructor's imagination is constrained by the actual fossil finds sparking that imagination, or free-ranging... ie how conservative or speculative their conceived dragon is. And some ideas might validly be entertained which are not strictly or solely derived from the fossil remains, but intuited otherwise, to be tested later.)
But using "dragon" could de-elitise the field of palaeontology and maybe make it even more appealing, universally accessible/welcoming across cultures and age-groups (mind you, kids already relish dinosaur names). People could call a thing they saw pictured or modelled 'a dragon' confidently, without embarrassment at not knowing its scientific (species or family) name. The more we discover about dinosaurs, it seems to me, the more (not less) they fit the archetypal dragon image. 'Dragon' is a wider-applicable term than 'dinosaur' (which itself is often colloquially coopted, misapplied to many non-dinosaurs eg plesiosaurs and pterosaurs and even Glyptodon, mammoths and more!). One could qualify it further and easily by habitat or stance: eg land dragons, sea dragons, sky-or-flying dragons; quadruped or biped dragons or flippered/finned dragons; all of which is pretty obvious at a glance.
One might still get Glyptodon or Doedicurus called 'a dragon' sometimes maybe by mistake. But, no system is perfect...!
Long live the dragons! Here's to dragons! Here be dragons!
:-D
Metalraptor:
Don't know much about some of those directly, but I've seen a lot of mekosuchine material. Except for Mekosuchus-Quinkana-Volia, other mekosuchines are ecomorphically within the range of modern crocodylids and alligatorids (or nearly so, for Baru). When Paul Willis first suggested Mekosuchus may have been a climber, I started paying attention to croc unguals from Riversleigh - and I haven't seen any that are laterally compressed or more strongly curved than in modern forms. Also, he based the idea partly on the 'fact' there were no varanids in Australia in the Oligo-Miocene, now known not to be the case. Postcrania of mekosuchins are still poorly known, but I've got most of a M. whitehunterensis skeleton here and the limbs and girdles are nothing extraordinary for a crocodylid, while the caudal neural spines show the tail was plenty deep enough for swimming. Quinkana goes fully xiphodont and may have been more terrestrial than Mekosuchus, but it was a better match for large terrestrial varanids (V. komodoensis to V. priscus-sized), with which it may have competed directly, than medium-sized arboreal ones.
Much longer comments occur all the time on Pharyngula. I've written several... Often the longest comments are in the longest threads, too.
That idea has been criticized, though unfortunately I don't know by heart where.
I said, again, the absolute height. Paleosuchus is tiny. If it held its head a meter above heart level (which of course it can't because its neck isn't that long), then you'd have a point.
That was the Nature editorial, not the paper itself. The paper just used the high rate of mitochondrial DNA evolution in crocodiles as evidence for a formerly higher metabolism (...which I don't find particularly convincing, actually).
I did equate the 1980s with "outdated", didn't I?
I'm talking about the last few years only. Even GSP hasn't published on the subject since 2002, AFAIK.
Of course I took it from Paul & Leahy 1994.
Growth -- turning amino acids into proteins -- is metabolism. Fast growth is fast metabolism. I'm not sure if it matters which comes first; the other will follow...
For each amino acid that's added to a growing protein, you need one molecule of ATP. That has to come from... a fast metabolism.
You can have fast metabolism without fast growth or even without any growth. But you can't have fast growth without fast metabolism.
No, DELTRAN is not more parsimonious than ACCTRAN.
We already know that the shift happened sometime between the origin of the total group and the crown-group of birds; all possible hypotheses on where between these two points it happened are, all else (like fossil evidence) being equal, equally parsimonious. Without looking at or in the bones, we have no default state that we could assume on this question.
In his talk at the SVP meeting in October, Unwin declared that he had been wrong about the articulation of the pteroid and Bennett had been almost right. The orientation and mobility he suggested in that talk were very close to those proposed by Bennett in his recent JVP paper.
Or is it two? One to charge the tRNA with the amino acid, one to move the ribosome forward? Or is it even more? I forgot... :-(
Jura
It seems very unlikely to me that anyone would assume so. Body shape and size of most of these, without (evidence for) insulating integumentary structures, make it pretty much impossible. Only the largest varanids at their highest latitudes (V. varius, rosenbergi and of course V. priscus in southeastern Oz, for example) might suggest something unusual among ectotherms. Ectotherms really can get bigger in the tropics (and the Naraccoorte and Wellington priscus material is all pretty small compared to the Darling Downs stuff, afaik).
It would also be hard or impossible to tell from fossils whether a snake was in the habit of cruising with the whole neck off the ground. Assuming a complete Naja skeleton in articulation, it's reasonable to assume an erect display could be inferred, but I think there'd be nothing so obvious in Demansia, Oxyuranus, Dendroaspis or even Ophiophagus (it's worth looking, but not easy).
How about the Svalbard light regime as a modern day example? Modern polar animals seem to manage without huge eyes, too (even if that's because of temperature constraints).
- Metalraptor wrote: That's why I call crocodilians, birds, non-avian dinosaurs, pterosaurs, and crurotarsians archosaurs rather than reptiles, and put them in a group on par with mammals.
Crurotarsi already includes Crocodylia. But that aside, the whole notion of putting one group "on par" with another sounds more like grades/ranks rather than clades. ;)
And besides, what about the lepidosaurs, turtles, and other non-archosaurians?!
Mark & David:
Erm, chameleons? Or do they only qualify as semi-erect?
Graham:
But that would lead to confusion with the several extant Australasian agamid 'dragons'. Therefore, alas, I vote 'Nay' on that suggestion.
Regarding the relation between neck-carriage and endothermy... It's perhaps better to wait for the soon-to-be-published study that Darren hinted at before, and get some actual data, before commenting further on that subject. But, despite the risk of repeating myself, I'd still like to remark on the following.
David:
Absolute head-above-heart height is the important factor? Fair enough, that makes sense. But that suggests the existence of some empirically discovered, measurable height threshold, above which ectotherms can't go but endotherms can. Are those kind of data available anywhere yet? (To me, holding the head a whole meter above heart level definitely sounds like an excessively harsh criterium. What extant mammals, apart from the giraffe, hold their heads that much higher than their hearts, anyway?)
Incidentally: while I'm not saying that what follows is necessarily relevant when discussing dinosaur metabolism, I'd like to point out that the ectothermic king cobra Ophiophagus hannah can reportedly hold its head, the neck fully erect, about one meter above ground. (Not one meter above its heart level, however, as the cobra's heart is situated quite far forward, in the raised part of the body.) And other cobras and certain other snakes can also hold their necks in a fully vertical position for quite some time, despite being ectotherms.
John:
Am I mistaken, or have there been claims to the effect that certain large Australian monitors (e.g., the perentie) are actually endotherms?
Dartian,
Camels and Llamas have their head around a meter or more above the heart, and Gerenuks and Dibatags when standing vertically probably do to.
I don't know if perenties have been considered endotherms but they use buccal pumps to get around carrier's constraint so they can be continuously active; selection might then lead to improvements in stance and then a higher metabolic rate.
LeeB.
I was so dang right.
LeeB:
I'm not so sure they do, at least not most of the time. Camelids have long necks, yes. But judging by, e.g., this picture of a dromedary in a fairly typical stance, the height difference between head and heart is normally considerably less than one meter. Llamas and the gazelle you mention perhaps typically hold their necks more fully erect than Camelus. But they're also much smaller animals, so I suspect the absolute distance between head & heart in these animals does not typically reach or exceed that 1 meter, either.
(And anyhow, all these mammals - giraffe, camelids, long-necked gazelle - are artiodactyls, so extreme long-neckedness can hardly be considered a very common feature among Mammalia.)
Thanks for the perentie info.
Oh yeah. I don't know by heart just how erect they can be, but they probably qualify. Let me explain it away by how tiny they all are. :-)
I don't know (and that's why I deliberately picked an almost certainly exaggerated threshold, apart from it being a nice round number). But it's probably possible to calculate the value from information that's already in the literature.
BTW, I forgot to comment on the wastefulness of tachymetaboly. Yes, it is wasteful, but it's an investment -- as long as food is available, it allows you to get even more energy than you lose!
Okay, I've heard of the "can't hold head more than 1 meter above the heart" argument before, and as others have already brought up, it seems kind of arbitrary. Especially since so many mammals with fairly long necks capable of being held a meter above their hearts (e.g. horses, wildebeest) spend so much of their time with their heads below the heart level (i.e.: http://www.wormsandgermsblog.com/Horse%20Grazing.jpg).
I thought Darren was referring to some other aspect of neck carriage I hadn't heard of.
Regarding fossil autarchoglossans, I suppose it would be more likely that they would just be thrown in the "'nother lizard" wastebin, but I'm now curious to see if there are some osteological correlates to things like maintenance of body posture in elapids. Looks like I'm going to have to get a hold of some snake skeletons now. :)
Sven wrote:
Glad to see I'm not the only one who sees a disconnect with that logic. It's also pertinent to note (and also a little strange that it never seems to get brought up) the amount of parental investment that mammals and birds devote to their young. Namely: they feed them. This is likely the main reason for higher growth rate in these two groups. Babies don't have to worry about where their next meal is coming from. The parents guarantee a steady supply of food so there is no need to store it as fat. Even captive raised, fast growing crocodylians, still convert 60% of the food they digest, into fat. Crocodiles could grow a heck of a lot faster if they didn't devote so much of their food to their "rainy day fund." It would be really interesting to test this statement by raising some croc hatchlings that have their fat deposit genes knocked out. I'm not sure what the ethics of that would be though.
David wrote:
Yeah, but that was already happening prior to the switch. Biomass conversion rates for bradymetabolic critters average around 50%, compared to ~1.5% for automatic endotherms. At the very least, it would not seem that energy acquisition was the main thrust for a tachymetabolic lifestyle.
Dartian wrote:
Varanids are capable of endogenous heat production (see: Bartholomew and Tucker, 1964 for more). They have been observed rhythmically "shivering" their muscles in order to warm the body, thus making them endothermic in the same sense as certain insects. As far as I know, that's the only association with endothermy that I've seen.
All this is true as far as it goes. Yes, growth is an energy-expensive process. The energy costs of growth are extremely important in the energy budgets of ectotherms; less so (because swamped by the insane costs of maintenance) in homeothermic endotherms. [See Peterson, C.C., B.M. Walton, and A.F. Bennett. 1999. Metabolic costs of growth in free-living garter snakes and the energy budgets of ectotherms. Functional Ecology 13:500-507.]
So let me refine my point. A fast-growing animal will have a high metabolic rate (=heat production) because it is growing fast. It must, therefore, have the physiological capacity to support the rate of energy expenditure due to growth.
BUT the claim that dinosaurs grew fast, therefore they were endothermic homeotherms goes beyond that, to positing an ineluctable relationship between high rates of growth (and concomitant high metabolic rate while growing) to tachymetabolism, which refers to the high resting metabolic rates that are necessary for bird/mammal-style endothermic homeothermy. It does matter which comes first, because growth is a pulsatile, not a constant, process, even in endotherms. That is the mechanistic link I am asking for. Without it, the argument confuses correlation for causation.
(By the way, the article cited above also documents that growth in herbivorous mammals is far less energy-efficient than in ectotherms.)
Megapodes.
It appears that it's not possible to elevate the metabolism above some multiple of the rest metabolism -- I forgot what multiple (and the citation of course). At least there's no known example.
Ah, the nugget of undemonstrated assumption at the core of the argument. Do the metabolic demands of high growth rates cause an ineluctable increase in maintenance costs exclusive of the actual processes of growth? If so, how (the increase) and why (ineluctable)?
This is the weak link.
"Ah, the nugget of undemonstrated assumption at the core of the argument." (Sven DiMilo) Really? A significant positive correlation has been demonstrated between BMR and growth rate in diverse animals and there is no indication that the mammal-like growth rates reported for dinosaurs can be sustained by ectotherms. The growth rates reported for dinosaurs seem best explained by endothermy, I am surprised that there is so much resistance to this logical inference.
A significant positive correlation has also been demonstrated between BMR and maximum exercise metabolism, IIRC.
No. Leaky cell membranes make the energy metabolism churn out so much ATP that the little extra for growth hardly matters.
Dr Jack wrote: "I am surprised that there is so much resistance to this logical inference." - in fact it isn't resistence, just that we are not all convinced it is a logical inference.
In the words of Sherlock Holmes "There is nothing more deceptive than an obvious fact" - it may seem obvious that features such as erect gait, erect head carriage and bone histology indicative of high consistent (i.e. non-seasonal) growth rate point to dinosaurs, and many other archosaurs, being truely endothermic, because all these tend to be associated with true endothermy today. But remember there are examples of extant ectotherms showing the above characteristics, and I still await any evidence that any of these features are functionally connected in any way to endothermy.
For example erect head carriage is about plumbing (a circulatory system that maintains sufficent blood pressure) rather than endothermy. Though most of the creatures today that have such circulatory systems are endotherms (and I can see why it would be advantageous), I am completely unconvinced that they are necessarily connected.
So I'm not resisting, I'm just not convinced. As I have already stated for pterosaurs and some theropods I find the evidence compelling, but for the rest I find it lacking. There is evidence for and against (such as lack of nasal turbinates and the energy claculations for some) - over all I guess if pressed I think the balance lies slightly in favour of endothermy for the other dinosaurs, but only just, and substantially in favour of ectothermy for the majority of non-ornithodiran archosaurs - but in reality the evidence is just not enough to draw meaningful conclusions.
And I'm with Mark on this. The argument from correlation at best generates a hypothesis; especially when the correlation is basically from only two data points: extant endotherms vs. extant ectotherms (maybe 3 given the independent evolution of endothermy in birds & mammals). I'm a skeptical physiologist--explain the mechanism and then I'll think about buying it.
Really?? You're going to have to show me those data. I try to pay attention to this stuff and I must have missed that.
Nah. The aerobic capacity model for the evolution of endothermy has been much discussed and only very poorly supported. If it works at all, it is because high aerobic capacity somehow drags up resting MR, NOT because a high RMR somehow powers exercise capacity. But any linkage is far from ineluctable...even among mammals alone, dogs have aerobic scopes approaching 30x and pronghorn antelopes 50x resting; most mammals can only do about 10x.
Mechanism, please.
Dr. Jack wrote:
I've read a lot of papers that have posited this claim, but very few that actually demonstrate it; especially if it involves reptiles (many refer to one, or two studies that give a blanket coverage of the topic).
There are definitely extant ectotherms that show growth rates on par, and in fact far exceeding, extant endotherms. Squid are quite possibly the fastest growing animals on the planet, with lifespans for even large species, rarely exceeding a few years (even the giant squid - Architeuthis dux- seems to live for only 6 years). Their growth is exponential and non-asymptotic (Jackson and O'Dor, 2001). Their growth rate is attributed to a high rate of food acquisition, and a very low (if nonexistent) allocation of that food to fat. Despite the fast growth, these animals are still temperature sensitive ectotherms (Forsythe, 2003).
David wrote:
I'm not sure where this initially came from, but I remember it was a central tenet in Bennet & Ruben's (1979) argument for endothermy through aerobic capacity. The central flaw in this theory still remains. Namely, why a high BMR, which is powered by the heart, kidneys, liver and intestines; should lead to increased aerobic capacity (powered by heart, lungs, and skeletal muscle). The argument simply seems to state that animals are only capable of increasing their active metabolism by 10x their resting, so a higher BMR should lead to a higher aerobic capacity.
20 years of metabolic research, seems to throw this idea in the trash though. As Sven wrote, dogs and antelope are capable of substantially higher aerobic scope than their BMR would suggest. Gila monsters (Heloderma) are able to maintain an active metabolism that is 30x that of their resting metabolism (Beck et al, 1995). While the greatest gap between BMR and active metabolism, comes from ectothermic flies (100x resting metabolism. Davis & Fraenkel, 1940).
Why would that be so? As mentioned before, leaky cell membranes incur a higher cost. Any extra ATP generated, is done in spite of the higher ATP requirements of those cells. There is no reason why a more costly metabolism should generate a net energy benefit. At least, I haven't seen a reason.
Refs:
______________
Beck, D.D., Dohm, M.R., Garland Jr, T., Ramirez-Bautista, A., Lowe, C.H. 1995. Locomotor Performance and Activity Energetics of Helodermatid Lizards. Copeia. Vol 3: 577-585.
Bennet, A., and Ruben, J. 1979. Endothermy and Activity in Vertebrates. Science. Vol. 206: 649-654.
Davis, R.A. and Fraenkel, G. 1940. The Oxygen Consumption of Flies During Flight. J.Exp.Biol. Vol 17: 402-407.
Forsythe, J.W. 2003. Accounting for the Effect of Temperature on Squid Growth in Nature: From Hypothesis to Practice. Marine & Freshwater Research. Vol 55(4): 331-339.
Jackson GD, OâDor RK (2001) Time, Space and Ecophysiology of Squid Growth, Life in the Fast Lane. Vie Milieu. Vol. 51: 205-215
Thanks, Jura, for the squid refs.
Certain moths can increase oxygen consumption while flying to 170x resting rates at the same environmental temperature (they are endothermic only while flying).
Positive correlation between BMR and growth rate does indeed appear valid, as shown by Whittow's data on juvenile birds, Coulson et al. on crocodilians, and in the data collected by Case. The high sustained growth rates reported for dinosaurs, well above those present in living reptiles, only have a parallel in endothermic birds and mammals. I think as soon as you start bringing altricial, thermally protected juvenile birds, giant squids and insects into the equation, you are avoiding the issue.Please explain how ectothermic reptiles might sustain mammal-like growth rates now seen in dinosaurs. We have no data indicating that this is possible, even in the most ideal of conditions. The endothermy hypothesis, while not conclusive, remains most parimonious.
Ah, but now we're moving the goal post. The argument, as I have seen it here and elsewhere, has always been that "endothermy" is a requisite for traits X,Y,Z, etc; and that "ectothermy" is incapable of providing the necessary power for said traits.
The qualifier: "in vertebrates" or "in a closed circulatory system" etc is never brought up. Apparently the alleged advantages of "endothermy" over "ectothermy" are supposed to be universal.
Well ectothermy and endothermy both cross numerous taxonomic lines; which seems to make practically all taxa fair game.
Well, that's something to work with; thanks. Specifics would be appreciated if handy.
I don't even mind restricting the dicussion to vertebrates or tetrapods; i just want to know how it's supposed to work. Mechanism.
Jura:
Whether parental feeding is the 'main reason' for rapid juvenile growth I am not qualified to say, but there seem to be at least some experimental data that support your suggestion (David's megapode counterexample notwithstanding). A few vertebrate ectotherms, mostly fish, do feed their young (see, e.g., Hildemann 1959; McKaye 1986). These include the convict cichlid Cichlasoma nigrofasciatum (Wisenden et al. 1995). And according to Zworykin et al. (2000), increased parental food provisioning does indeed lead to faster growth rates in the fry of this species.
References:
Hildemann, W.H. 1959. A cichlid fish, Symphysodon discus, with unique nurture habits. The American Naturalist 93, 27-34.
McKaye, K.R. 1986. Trophic eggs and parental foraging for young by the catfish Bagrus meridionalis of Lake Malawi, Africa. Oecologia 69, 367-369.
Wisenden, B.D., Lanfranconi-Izawa, T.L. & Keenleyside, M.H.A. 1995. Fin digging and leaf lifting by the convict cichlid Cichlasoma nigrofasciatum: examples of parental food provisioning. Animal Behaviour 49, 623-631.
Zworykin, D.D., Budaev, S.V. & Mochek, A.D. 2000. Does parental fin digging improve feeding opportunities for offspring in the convict cichlid? Environmental Biology of Fishes 57, 443-449.
For a Brachiosaurus or Giraffatitan with an erect neck, Paul & Leahy (1994) calculated that a heart capable of generating such high pressures would alone have a higher metabolism than an entire sauropod-sized bradymetabolic animal. Granted, this says nothing about most dinosaurs, but it does make bradymetabolic sauropods impossible.
(Yes, this does require that Stevens & Parrish are wrong about the habitual neck position of Giraffatitan. And indeed they are wrong -- H. Mallison, pers. comm., citing his own extensive but unpublished work on sauropodomorph biomechanics.)
It has long been known, and published, that ossified (!!!) nasal turbinates were in fact present in several Mesozoic dinosaurs. Isn't there a paper in the Ostrom Symposium volume (2001) on that?
OK.
Not endothermy, but tachymetaboly. Growing is hard work.
Yes, but -- compared to vertebrates -- they cheat: they add new muscle cells throughout their lifetimes, something vertebrates just don't do.
If that's not enough, I suppose the lack of hard parts probably also helps.
To repeat myself, it's an investment. It allows you to get so much food that you can more than make up for your waste.
(If, that is, enough food is available a priori. In some environments it isn't, and that's why tachymetabolic animals are absent or restricted there.)
If there is no net energy benefit to a more costly metabolism, why do animals with a costly metabolism exist at all? Why isn't automatic tachymetaboly selected against in all environments?
No, it's simply understood that we're comparing crocodiles, lepidosaurs, turtles and maybe lissamphibians to ornithodirans and mammals -- so it doesn't need to be mentioned all the time.
How much faster?
David:
Annoyingly, the authors do not tell that! They state quite clearly that juvenile cichlids grow faster the more frequently the parents provide food for them (there is much individual variation in parental care intensity in this species) but they don't give the raw data for growth rates! I suppose those data do exist, though, so if someone is really interested, it's probably better to contact the authors directly.
David wrote:
Are you sure about that (seems it would have made big news on the DML if that was the case)? Are you sure that they weren't talking about rostral turbinates (which are found in various dinos like Nanotyrannus, as well as many extant reptiles)?
That's the $64,000 question. Automatic endothermy (endothermy through a tachymetabolism) is very costly, and the benefits are not as obvious as was once thought. This is part of the reason why it has been so hard to figure out how it could have even evolved in the first place. Net energy benefit seems unlikely since, compared to extant ectotherms, automatic endotherms are actually taking a net energy cut. Being a constant temperature all the time seems like a more likely scenario (though the "hot is good" theory is currently out of favour based on the current evidence). In short: we don't know.
RE: squid
I don't doubt that both are likely contributors. Regardless, they do still show that ectothermy, by itself, is not a hinderance to speedy growth.
In that regard:
But it's not even mentioned some of the time. The argument is always one of "endothermy" vs "ectothermy." There is never a mention of taxa limitation.
Dartian wrote:
Adding to that list is the rather recent discovery of parental feeding in some caecilians (Kupfer et al, 2006)
Sadly, not much seems to be written besides a description of the act. The authors do mention that the young are able to increase their birth weight by an order of magnitude at the end of their first week. That's certainly no mean feat.
Finally, the growth rate for megapodes is remarkably slow (for a bird). Mallee fowl (the only megapode that appear to ahve actual data on this) show a growth rate of about 0.01g/day (Starck & Ricklefs, 1998). If one were to insist on using Case's 1978 study, then it would seem to throw Mallee fowl square in the "reptile" camp.
Refs:
_____________
Kupfer, A., Muller, H., Antoniazzi, M.M., Jared C., Greven, H., Nussbaum, R.A., Wilkinson, M. 2006. Parental Investment by Skin Feeding in a Caecilian Amphibian. Nature. Vol. 440(7086): 926â929.
Starck, J. and Ricklefs, R.E. 1998. Avian Growth and Development. Oxford U. Press. ISBN: 0195106083. page: 250.
Yes, and specifically respiratory as opposed to olfactory turbinates, as distinguished by their position in the nasal cavity. I don't have my literature here in Paris, but I think GSP's chapter in the Ostrom Symposium volume talks about that at some length, and if not, DA does.
Then what do you think of the investment hypothesis? As in, in a generally food-rich environment, being constantly active allows you to get so much energy that -- in spite of all the waste -- you still have (in absolute terms!) more energy left that you can invest in reproduction?
That was just the missing link between having free-living aquatic larvae and having the young inside the oviducts and eating the oviduct lining. No more surprising than all the viviparous frogs, salamanders, and squamates.
Now this is interesting!
Dartian:
Cichlasoma was one horribly paraphyletic genus in its heyday - it has since exploded into numerous genera. The convict cichlid is now Amatitlania nigrofasciata.
David:
Did you mean that squids have a relative lack of hard parts? They possess a shell remnant composed of calcium carbonate or chitin, a chitinous beak, and vertebrate-like hyaline cartilage in the cranium and other structures. I haven't come across any readily available figures for how much this all weighs, but since the arms and tentacles lack hard structures it seems reasonable to assume that their hard parts amount to considerably less than a vertebrates' proportionally. I didn't know about the continuous addition of muscle cells, fascinating!
So does this count as off topic?
Cichlid taxonomy and squid morphology aren't off-topic, given that those subjects were arrived at by logical succession :) I think this comment I'm writing now makes it 98 for this post - who wants to make it to 100? You're not allowed to cheat and post nonsense, however.
Nasal turbinates: the hypothesis, such as it is, that these are required for endothermy is suspect given that some extant endotherms lack them. And, yes, note that 'RT are correlated with endothermy' is itself a hypothesis, as no-one has ever demonstrated it. Because we can't be sure whether Mesozoic archosaurs had RT or not (due to the vagaries of preservation), Ruben et al. (1996, 2003) attempted to reconstruct the size and extent of the nasal passages. They concluded that the nasal passages of Mesozoic archosaurs were too small for RT, and hence that RT were absent. However, they cheated, both by mis-reconstructing the size of the nasal passages, and by mis-reconstructing the path of airflow through dinosaur and pterosaur snouts. Greg Paul later argued that dinosaurs may well have possessed RTs, based on the size of the nasal cavities: there's a whole appendix dedicated to this subject in Dinosaurs of the Air. It has indeed been claimed on occasion that RTs have been discovered in dinosaurs - I'm pretty sure Larry Witmer said this at one of the SVP meetings - but the turbinates discovered so far all seem to be olfactory (see Witmer & Ridgely 2008 for ankylosaurs).
Refs - -
Ruben, J., Hillenius, W., Geist, N. R., Leitch, A., Jones, T. D., Currie, P. J., Horner, J. R. & Espe, G. 1996. The metabolic status of some Late Cretaceous dinosaurs. Science 273, 1204-1207.
- ., Jones, T. D. & Geist, N. R. 2003. Respiratory and reproductive paleophysiology of dinosaurs and early birds. Physiological and Biochemical Zoology 76, 141-164.
Witmer, L. M. & Ridgely, A. C. 2008. The paranasal air sinuses of predatory and armored dinosaurs (Archosauria: Theropoda and Ankylosauria) and their contribution to cephalic structure. The Anatomical Record 291, 1362â1388.
Great discussion.
Agreed that endothermy does not require respiratory turbinates. However, from a functional standpoint, we know how RTs function to conserve water and heat in extant endotherms that have them, and not only do no extant ectotherms have them, but nobody has yet suggested a plausible reason why they should. I regard the presence of RTs as a strong indicator of tachymetabolic endothermy (same with plumage/pelage insulation), but their absence means diddly.
Jura is correct that the evolutionary benefits of tachymetabolic endothermy are still far from clear. A whole issue of Physiological and Biochemical Zoology (2004, vol 77(6)) was recently dedicated to the topic. Lots of interesting reading in there. Of all the scenarios presented so far, I like Koteja's ideas best (pdf available here).
Constant body temperature seems like a great reason to me. To survive in colder climates (like here, in Alaska), animals need high metabolisms and constant, high body temperatures. And many animals still hibernate--both ectothermic (wood frogs) and endothermic (bears). However, I imagine bears hibernate because the metabolic costs of operating in the depths of winter would outweigh the potential food sources.
Bears don't just eat meat, after all. They eat berries and fish (technically meat, I suppose), both of which are absent in the winter. Moose wander around, but unless it's a juvenile moose without a mother, any bear would be a fool to try and take it down. So I imagine bears hibernate because their food options are so limited.
Moose eat all year long, though. They're quite adept at surviving on low-quality winter vegetation.
Mallee fowl (the only megapode that appear to ahve actual data on this) show a growth rate of about 0.01g/day (Starck & Ricklefs, 1998).
I don't question that megapodes might grow slowly, but the cited rate of one one-hundredth of a gram per day seems unlikely. At that rate it would take 100 days to add a single gram, and over a century to put on a pound. Is it possible that either the decimal is in the wrong place or the unit is wrong? I ask because I don't have the reference handy, and I would really like to know the exact number. Thanks in advance.
>Dr. Vector
You can access (most of) the book here.
Re: megapode growth rates, check out these papers...
Göth, A. & Evans, C. S. 2004. Egg size predicts motor performance and postnatal weight gain of Australian Brush-turkey (Alectura lathami) hatchlings. Canadian Journal of Zoology 82, 972-97.
Stark, J. M. & Sutter, E. 2003. Patterns of growth and heterochrony in moundbuilders (Megapodiidae) and fowl (Phasianidae). Journal of Avian Biology 31, 527-547.
Göth & Evans (2004) - I have a pdf if anyone wants it - includes growth curve data for brush-turkey hatchlings. At hatching, mass is between 90-160g; the growth data then shows them reaching 500g by 2 months old, and 1000g by 3 months old (fig. 5, p. 976). An increase of 500g in c. 30 days amounts to c. 17g per day, somewhat higher than the figure mentioned above! I don't think these growth rates are much different from those of other galliforms.
Squid? As opposed to cuttlefish?
Organic matter.
(Though probably not cheap either. But then, it's not a whole skeleton.)
But there were no colder climates in the Triassic!
I checked the paper again, and that is what it said. It's still a bit strange. For instance, it says .01 /day. No actual value is there, despite the fact that their growth rate equation shows adult weight measured in grams (almost wonder if it's a typo).
I did, however, find a different paper with better data in it.
Gillespie, J. Captive Breeding and Management of Mallee Fowl (Leiopoa ocellata) at Western Plains Zoo, 1989 to Present (pdf available here
Mallee fowl are born at 112g hatching weight, and grow at a rate of 2-3g per day. Adults weight is 2kg, which would put skeletal maturity at 2.6 years for the low end. This doesn't count the fact that the initial week, or two of a mallee fowl (and other megapodes) life is followed by a decrease in hatching weight.
It does however agree with previous studies (Jones et al, 1995), which state skeletal maturity to occur at around 2-3 years of age. That would appear to put them in Case's reptile graph (which also has a few wonky numbers in it. For instance: an 80kg Boa constrictor taking 91 years to reach adult size).
Refs:
______________
Gillespie, J. Captive Breeding and Management of Mallee Fowl (Leiopoa ocellata) at Western Plains Zoo, 1989 to Present
Jones, D.N., Dekker, R.W., Roselaar, C.S., van Perlo, B. 1995. The Megapodes. Oxford U. Press. ISBN: 9780198546511
[from Darren: delayed by spam filter, sigh]
The closest appropriate article... So, Daren, I hope you'll like it:
http://www.sivatherium.h12.ru/library/Dixon_2/00_en.htm
It is the second D. Dixon's book. The translation into Russian will be too hard because of too strange names of his critters.
The squid pen is chitinous.
David:
The common usage of "squid" includes every decapodiform which isn't a cuttlefish. Despite being the probable sister group of cuttlefish (or at least a closely related sepioid) Spirula is often referred to as a squid and is commonly known as the ram's horn squid. It does possess a calcareous shell. And this is why I avoid the term "squid" as much as possible...
Hmm, strange that it didn't post the first time. I must be misspelling my e-mail, or something. [from Darren: for some reason your comments are getting held up as spam. It might be because your comments include more than one url (your own site url, and the one you cite). I publish them as soon as I see them.]
Repost
_______________________________________________
I checked the paper again, and that is what it said. It's still a bit strange. For instance, it says .01 /day. No actual value is there, despite the fact that their growth rate equation shows adult weight measured in grams (almost wonder if it's a typo).
I did, however, find a different paper with better data in it.
Gillespie, J. Captive Breeding and Management of Mallee Fowl (Leiopoa ocellata) at Western Plains Zoo, 1989 to Present (pdf available here
Mallee fowl are born at 112g hatching weight, and grow at a rate of 2-3g per day. Adults weight is 2kg, which would put skeletal maturity at 2.6 years for the low end. This doesn't count the fact that the initial week, or two of a mallee fowl (and other megapodes) life is followed by a decrease in hatching weight.
It does however agree with previous studies (Jones et al, 1995), which state skeletal maturity to occur at around 2-3 years of age. That would appear to put them in Case's reptile graph (which also has a few wonky numbers in it. For instance: an 80kg Boa constrictor taking 91 years to reach adult size).
Refs:
______________
Gillespie, J. Captive Breeding and Management of Mallee Fowl (Leiopoa ocellata) at Western Plains Zoo, 1989 to Present
Jones, D.N., Dekker, R.W., Roselaar, C.S., van Perlo, B. 1995. The Megapodes. Oxford U. Press. ISBN: 9780198546511
Wow, we've reached a three-digit number of comments. Darren, you have created a monster!
(Just kidding. This thread is brilliant.)
Cameron:
Thanks for the information. It's good that Cichlasoma has been chopped up into something more manageable - although it was never as bad as that ultimate wastebasket fish genus, Barbus.
Zach:
Actually, I think it's rather the other way around. Brown/grizzly bears and black bears don't hibernate* because they have to, but because they can. They can afford to build up sufficient fat reserves thanks to their varied, high-energy summertime diet. (On the other hand, the giant panda, which has more limited food options than any other bear, does not hibernate; it can't build up large enough fat reserves to last through the entire winter.) Hibernation shouldn't be thought of as some kind of inferior survival strategy. If anything, it is probably a much safer and less costly way to get through the winter than staying active or migrating south.
* Strictly speaking, bears and certain other omnivorous carnivores - raccoons, Eurasian badgers, skunks, raccoon dogs - don't hibernate in the same way as ectotherms or certain smaller mammals (bats, hedgehogs, marmots, dormice, etc.) do. Their torpor is much less deep, and they wake up more easily and regularily.
It's probably expressed as a specific growth rate, i.e. g/(g d).
I've seen the term "carnivoran lethargy" used for the shallow "hibernation" of bears et al.
Note, btw, that the capacity for true torpor may well be ancestral for therians.
German has a separate term for it: Winterruhe "winter rest", as opposed to Winterschlaf "winter sleep", which is true marmot-style hibernation. Zoologists have long insisted on the distinction.
(Erm, just to be clear, zoologists have also invented the distinction. Winterruhe is a neologism that can't possibly be more than 300 years old, likely less than half that.)
Darren, why does this fantastic blog post not link to ResearchBlogging?! I almost missed it, if it hadn't been for my brother-in-law. Best of luck!
A thought-provoking thread of commentary. I'm a biochemist, not an evolutionary biologist and certainly not a dinosaur afficianado; so my perspective is admittedly naive. But a few questions arise which ought to be answerable experimentally:
1. Have genes been identified which are necessary for endothermy via the ion-permeable membrane mechanism of heat production? Are these genes carried by the autosomes or by the mitochondrial DNA?
2. Have any studies been carried out on the crocodilian genome to determine whether any of these endothermy genes are present as pseudogenes or unexpressed coding domains?
If the hypothesis is correct that modern crocs descend from endothermic ancestors, then the traces of that ancestry may linger in their genome: mutations which silence genes are more common than those which delete them entirely.
3. If genes necessary for endothermy have been identified, what does their mutation rate over time, as inferred from sequence divergence in lineages (mammals vs birds vs reptiles) suggest about the antiquity of these genes?
4. Does the shift from exotherm to endotherm impose a selection regime on mitochondria? This seems a very fundamental metabolic question, but I must admit I never thought about it before. If it does - and offhand I suppose it must - then traces of that evolutionary selection should reside in the mitochondrial genome just as much as in the autosomes of the host. Mitochondrial DNA tracing of the maternal lineage through mutations in noncoding regions is well established. Functional mutations in coding regions to enhance endothermy should extend the time window for tracing mitochondrial lineages.
I smell a PhD thesis.
No, not to my knowledge. The cellular determinants of metabolic rate are understood at all well only for mice, rats, and one or two lizard species. Birds are poorly studied, afaik (correction solicited from anybody who knows better!) There's a lot going on. Rodents vs. same-sized lizards have: larger energy-intensive organs (liver, brain, heart, gut); higher rates of protein synthesis (whole-animal, tissue, and cell levels); leakier cell membranes to ions and concomitant higher ion-pump activity; and leakier inner mitochondrial membranes to protons (this latter results in markedly less efficient ATP production, and its function, beyond heat production, is debated). On the supply side, rodents have higher densities of mitochondria in tissues, and increased inner-mitochondrial-membrane surface-area and aerobic enzyme activity.
I'm sure somebody is working on microarray studies to identify candidate genes, but I haven't seen any results yet. One hypothesis is that the membrane-based differences are mediated by phospholipid composition (search Else & Hulbert, or see the issue of PBZ I cited above); this would suggest enzymes expressed in the smooth endoplasmic reticulum...
2., 3. Great ideas! Someday, maybe...
4. Also a cool idea. Unfortunately (again, afaik), the genetic controls of mitochondrial density and membrane SA are unknown, and (it seems to me) quite likely to be nuclear.
[Just found a table (Else, Turner & Hulbert 2004; PBZ 77:951) that compares mitochondria data for hearts and livers of five small endotherms (sparrow, bat, mouse, K-rat, dasyurid) to four ectotherms (toad, 2 lizards, garter snake). Endotherms with higher densities of mitochondria and inner membranes; sparrows not different from mammals but emphasizing liver over heart.]
Mallee fowl are born at 112g hatching weight, and grow at a rate of 2-3g per day. Adults weight is 2kg, which would put skeletal maturity at 2.6 years for the low end....It does however agree with previous studies (Jones et al, 1995), which state skeletal maturity to occur at around 2-3 years of age.
Many thanks for the follow-up information and citations. I was under the (evidently mistaken) impression that extant birds get to full size in a year or less, so they aren't growing for long enough to form annual LAGs. Has anyone done any bone histo on megapodes?
Just a bunch of questions:
I did not know that a prepubis was known for Anhanguera and yet one was illustrated in the Classens, O'Connor, Unwin paper. Any refs you can point me toward?
I've always been under the impression that the prepubis/pubis joint was a butt joint permitting very little movement. Any good refs on that connection?
I also wondered about the role of size and shape of the prepubis. In some taxa the prepubis is much deeper than in others. The shapes vary with each taxon and vary widely within Rhamphorhynchus, for instance. In the article there was a homogeneity that might have left another impression.
And finally I wondered about the size of the pubis, which can be extremely short in certain taxa, much shorter than the ischium. So the prepubis in these cases attaches much closer to the acetabulum. I wondered how this might have affected the running hypothesis?
Hi David - I think you need info from the authors regarding these questions (I'm meeting up with Unwin tomorrow, actually, and will ask him then). However...
-- An Anhanguera prepubis is indeed not figured by Wellnhofer or Kellner & Tomida, but Claessens et al. only figure one in their soft-tissue reconstruction, and I presume that this incorporates data from other ornithocheiroid taxa (such as Pteranodon).
-- Movement of the prepubes: the articular surface of the prepubis was described by Bennett as concave and cylindrical in Pteranodon, and I think this is the main reason for thinking that the prepubes were dorsoventrally mobile. Note also that Claessen's work on the mobile crocodilian pubis provides a model for prepubis function in pterosaurs (Claessens 2004).
-- Prepubes vary in shape among pterosaurs - look at fig. 5 in Sayão & Kellner (2006) - but are always paddle-shaped or Y-shaped elements, connected medially, that bridge the gap between the pubes and gastralia. It seems reasonable to think that they all functioned in the same way, even if they vary in length as you note.
Ref - -
Claessens, L. P. A. M. 2004. Archosaurian respiration and the pelvic girdle aspiration breathing of crocodyliforms. Proceedings of the Royal Society London B 271, 1461-1465.
Sayão, J. M. & Kellner, A. W. A. 2006. Novo esqueleto parcial de pterossauro (Pterodactyloidea, Tapejaridae) do Membro Crato (Aptiano), Formação Santana, Bacia do Ararripe, nordeste do Brasil. Estudos Geológicos 16, 16-40.
I used to support endothermic dinosaurs; I've since joined Jura's ectothermic/everything's still equivocal camp. For now I'll just mention that after Farmer and Sanders' shiny new work on alligator lungs, the argument that unidirectional air flow/presence of air sacs indicates endothermy is totally moot and can be declared DOTS (Dead On The Spot, a term I just made up, and that will never catch on :)). The growth rate arguments are likewise screwy and suspect IMHO.
I also personally find it odd (read: unlikely) that dinosaurs would have been under selective pressure to evolve endothermy in the hothouse enviroments of the Mesozoic (I'm specifically talking about the Tiassic, when the transition should have occurred).
Tsk, tsk, tsk. First, it's not on the same level as in birds or sauropods; second, it's not clear what kind of metabolism the earliest crocodylomorphs actually had. All this stuff is probably gradual.
Sounds like an argument from personal incredulity to me.
It must have happened during the origin of ornithodirans (or even crown-group archosaurs) and for greater aerobic endurance.
Mammal endothermy evolved for staying warm. Bird endothermy evolved for keeping running. There was a paper recently that almost used these words, unfortunately I don't have it.
See, however, this paper (pdf) and the discussion section of this one. Google Scholar is your friend.
"Tsk, tsk, tsk. First, it's not on the same level as in birds or sauropods; second, it's not clear what kind of metabolism the earliest crocodylomorphs actually had. All this stuff is probably gradual."
It might not be on the same level as theropods or sauropods, sure, but the point is that Farmer and Sanders' work reveals that unidirectional air flow is not inevitably linked with endothermy, as had been implied and/or stated previously. It certianly raises the possibility that this could go one small step farther, with an ectothermic animal nevertheless having air sacs. And it still destroys the direct link between flow-through lungs and endothermy.
"Sounds like an argument from personal incredulity to me"
No, I don't think it is: The point is not that I think the growh rate studies are wrong, per se, it is the arguments that these growth rates have to indicate endothermy that I find suspect. Not the growth rates themselves, the pro-endothermy arguments founded on them. And anyway, it turns out sauropods didn't grow at the lightning-fast rates we thought they did (Lehman and Woodward 2008). It's also well known that crocodilians raised under optimal conditions grow at zippity-quick rates comparable to those of endotherms (sorry I don't have an actual ref on me right now). A big component in these 'optimal conditions' is the handing out of free food by humans; without having to actively search for prey (or evade predators for that matter), the hatchling can devote much more of its energy to growth and acheive higher growth rates, twice if not more of what it can acheive in the wild. We know that altricial hadrosaur hatchlings did little more than sit in nests all day long being stuffed with free food by the adults; substantial amounts of energy would therfore get devoted solely to growth, and high growth rates would probably have been inevitable regardless of metabolic status. Same goes for big theropods, which also seem to have fed their young (Bakker 1997; and yes, I am aware that there is a bit of controversy surounding this report).
And I screwed up on the hot environment-endothermy thing; I was very sleepy, as could probably be desiphered due to my misspelling of Triassic as "Tiassic" :)
Refs:
Bakker, Robert T. (1997). Raptor Family values: Allosaur parents brought giant carcasses into their lair to feed their young. in Wolberg, Donald L.; Sump, Edmund; and Rosenberg, Gary D. (eds.). Dinofest International, Proceedings of a Symposium Held at Arizona State.
Lehman, T.M., and Woodward, H.N. 2008. Modeling Growth Rates for Sauropod Dinosaurs. Paleobiology. Vol. 34(2): 264-281.
Could pterosaurs and birds have a common ancestor? There seem to be a lot of similarities.
Sure they do. Pterosaurs are ornithodirans that branched off before dinosaurs, which includes birds. However, a great many of their similarities are probably convergences, including their skeletal pneumaticity, as ornithischians do NOT have many of the birdlike features found in pterosaurs and maniraptors.
Can you list the characteristics that pterosaurs have that basal birds do not have? I know about feathers.
I should have said
Can you list the characteristics that pterosaurs have that modern birds do not have? I know about feathers.
And conversely what characteristics do modern birds have that pterosaurs did not have. Again I know about feathers.
I don't understand what you're asking for. I mean, even a quick glance at a pterosaur skeleton compared to a bird skeleton (basal or modern) should answer your question.
Zachary does not understand what I am asking for.
Can anyone else answer my questions?
Come on. Wing membranes, for starters!
Everything associated with them, means, a lot of forelimb and hindlimb anatomy!
Better yet, the finger that supports more than half of a pterosaur wing is the 4th, and that finger is completely missing in birds.
Teeth (completely irrelevant, but true).
Lack of clavicles (collarbones). In birds, they're fused to each other and form the furcula (wishbone).
Gastralia (again completely irrelevant, but true).
The bones between the eye socket and the temporal fenestrae (...again completely irrelevant, but true).
And I could go on for hours.
Really, what purpose does your question serve? Do you want to know derived character states present in pterosaurs but absent in birds? Do you want to know derived character states present in birds and their closest relatives but not in pterosaurs? I'm not sure you understand your own question. :-)
Jack: pterosaurs and birds are different in just about all detailed aspects of their anatomy - birds are theropod dinosaurs and hence possess numerous anatomical characters typical of theropods and dinosaurs as a whole, while pterosaurs are not dinosaurs and possess a different basic set of features. As Zach said, the differences are many and obvious. Nevertheless, here are a basic list of key pterosaurian and bird features - I've concentrated on the features present in the earliest members of the two groups.
_A few basic features present in pterosaurs not seen in birds_
-- Four-fingered hand with hypertrophied 4th finger forming main wing spar
-- Wing membranes stretching between wing skeleton and body and hind limb
-- Hypertrophied 4th metacarpal with twisted, roller-like distal end
-- Pteroid bone supports propatagium
-- Ilium slender and shallow dorsoventrally
-- Pubis and ischium short
-- Prepubic bones in front of pelvis
-- Five-toed foot with long 5th toe
_A few basic features present in birds not seen in pterosaurs_
-- Three-fingered hand with elongate fingers
-- Three metacarpals, with first mc much shorter than second and third
-- Semilunate carpal bone in wrist
-- Furcula (wishbone)
-- Ilium relatively deep, with expanded preacetabular region
-- Pubis and ischium long
-- Four-toed foot with first toe (hallux) shorter than other three
Pterosaurs and birds do share an ancestor, but that ancestor was a Triassic archosaur that gave rise to all ornithodirans: Ornithodira = the dinosaur-pterosaur clade. Any similarities present between pterosaurs and birds (and, apart from features that are widespread within Ornithodira, wings are about the only ones) are obvious convergences.
Jack: Go here.
It would be helpful to look at each one of these differences individually. I am hoping this will not just become an argument. It truly is a fascinating subject.
Pterosaur:
"Four-fingered hand with hypertrophied 4th finger forming main wing spar."
http://www.plosone.org/article/info:doi/10.1371/journal.pone.0002271
"Morphological features common to all azhdarchids include a long, shallow rostrum; elongate, cylindrical cervical vertebrae that formed a long and unusually inflexible neck; and proportionally short wings with an abbreviated fourth phalanx."
We see a development of the "4th" finger being shortened in advanced pterosaurs, becoming more bird-like.
In fact, as it becomes more bird-like, some of the "fingers" fuse as seen in modern birds.
(I will have more to say about this "4th" finger".)
Ok... so, you're implying that there might be a direct phylogenetic link between pterosaurs and birds. Good luck with that :) The paper you link to above is by Witton & Naish: I'm the 'Naish' if you didn't know. Azhdarchoid pterosaurs do indeed have an abbreviated fourth phalanx - in fact, some pterosaurs lacked the fourth phalanx whatsoever - but if you think this makes their wings in the least bit bird-like: well, think again. And pterosaur fingers do not fuse as they do in birds. Pterosaurs are a bit similar to birds thanks to convergence, but birds are theropod dinosaurs - I suggest you read the literature on bird origins and theropod anatomy.
Pterosaur:
"Teeth (completely irrelevant, but true)."
http://en.wikipedia.org/wiki/Pterodactyloidea
"The most advanced forms also lack teeth."
The more advanced pterosaurs are more bird-like.
I did know that you were the Naish. (Gives it credibility don't you think?).
I have read the literature.
I am hoping to discuss each difference and get people's thoughts.
I appreciate your wish of "good luck" but it is more a matter of following the evidence where it leads, rather than luck, as I see it.
I will turn to the subject of the number of fingers next.
The pterosaur has five fingers.
The pteroid bone is the evolved form of the thumb. (I know there is some differing opinions about which way it pointed).
The so-called "4th" finger is actually the 5th finger.
With this idea, there is now no problem in lining up the fingers and no need for complicated explanations like "frame shift".
No fingers have been lost.
Obviously the thumb has become a more fused structure since it is now being used to control the propatagium.
In modern birds this "thumb" is the alula.
Concerning the number of toes. Some pterosaurs had 4-toed feet, like birds.
http://en.wikipedia.org/wiki/Pterosaur
"However, a large number of pterosaur trackways were later found with a distinctive four-toed hind foot >/b>and three-toed front foot."
http://blog.everythingdinosaur.co.uk/blog/_archives/2008/2/16/3526757.h…
"The animal has been named Nemicolopterus crypticus, the fossil shows a number of adaptations for a life amongst the tress. For example, the eye sockets are quite large, indicating that this Pterosaur could have coped well with low light levels in the understorey of a dense primeval forest. The four-toed hind feet had sharp claws, which could possibly have been able to grasp branches, giving this tiny flying reptile a good purchase amongst the tree tops."
Jack, the pteroid bone is a part of the wrist. The thumb is in fact lost in even the most basal pterosaurs. Nemicolopterus is probably a juvenile of something else. Also, note that pterosaurs are plantigrade--they put the whole foot on the ground, whereas birds are digigrade--they walk tippie-toe. Also pterosaurs don't have a reversed hallux.
Birds don't have pteroid bones, either.
Jack: please don't take this the wrong way, but I don't appreciate you turning the comments section into a list of personal musings. The bottom line is that, even when superficial (note: superficial) features shared by pterosaurs and birds look alike, a better understanding shows that these similarities have _clearly_ arisen by convergence. If pterosaurs and birds were the only archosaurs around, then I can understand that postulating a close link between them would appear viable. But there are hundreds of other taxa to take into account: birds share innumerable characters with theropods, with other saurischians, and with other dinosaurs that are not present in pterosaurs. You need to take this on board; at the moment you are being misled by a few extremely superficial similarities.
I did a quick google on the phrase"
"the pteroid bone is a part of the wrist" and found this:
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1560000/
"The pteroid bone is a rod-like element found only in pterosaurs, the flying reptiles of the Mesozoic. It articulated at the wrist, and supported a membranous forewing in front of the inner part of the wing spar."
It would appear that it is not part of the wrist but "articulated at the wrist".
The most parsimonious idea is that it is an evolved thumb.
Is there some way to definitively disprove that idea?
"birds share innumerable characters with theropods, with other saurischians, and with other dinosaurs that are not present in pterosaurs. You need to take this on board; at the moment you are being misled by a few extremely superficial similarities."
I have not even listed the similarities yet. I have been addressing the specific "differences" that people have posted. And I have shown that there is a very straightforward explanation of the items I have addressed that you and David MarjanoviÄ listed. And I am desirous to address the others.
If you wish to stop discussion of this topic that is fine.
But I am very interested in the specifics of this.
It may well be that there are "innumerable" differences but that has definitely not been shown yet.
It may be a good time to mention that Jack has some history with me and Dave Hone. He attempted to argue that pterosaurs actually evolved from BATS over email with me, until I mentioned that pterosaurs were long dead before bats came around, at which point our man decided that I was right, and that bats evolved from pterosaurs. I reminded Jack that bats are mammals while pterosaurs are reptiles. He consented, sort of, and now argued that pterosaurs evolved into birds. Dave Hone had similar rambling nonesensical arguments with the man, to no end.
For whatever reason, Jack very much would like to believe that the entire paleontological community is wrong, and that birds evolved from pterosaurs. Why, I can't be certain, as he seemed equally happy with the idea that pterosaurs evolved from bats, and vice versa. However, he seems distinctly unhappy with the general consensus, well supported, that birds evolved from maniraptor theropod dinosaurs. The morphological differences between, say, Velociraptor and Archaeopteryx are virtually nil by comparison to the differences between Archaeopteryx (or even Velociraptor) and Pterodactyl.
Zachary has misrepresented the situation.
But I am not looking for an argument.
I am interested in the facts of the matter.
But it is always easier for people to personalize the discussion instead of dealing with the issues.
I have addressed most of the issues that people raised.
I will discuss the other issues if there is interest.
In fact, you have not addressed anything. You've misinterpreted and cherry-picked. That is not the same.
And actually, yes, I do have one issue of interest I'd like you to address:
After fully digesting the literature on pterosaur skeletal anatomy so that you can actually make a case for this, explain to me why pterosaurs are a better model for the avian ancestor that, say, raptor dinosaurs, which have feathers, by the way, while pterosaurs do not.
If paleontology has already unearthed a suitable avian ancestor, why cock it up by insisting that a completely unrelated group of reptiles provides a BETTER ancestry?
Jack, "phalanx" does not mean "finger", it means "finger bone". Each of your fingers and toes has 3 phalanges, except for your thumbs and big toes, which have only 2.
Well... no. They're not fused in any pterosaurs, or for that matter in any birds!
So what? Teeth were lost twice independently within pterosaurs and at the very least 3 times independently within birds. Ancestrally, the jaws of both pterosaurs and birds are lined with teeth.
The oldest toothless pterosaurs are younger than the oldest birds...
What makes you think so?
Check out the way it looks, and the place where it articulates with the wrist.
There were till 2008.
Dude... no such explanation was ever necessary. The normal phalangeal formula for amniote hands is 2-3-4-5-3 (2 phalanges in the thumb, 3 in the index finger, and so on). Dinosaurs have at most 2-3-4-3-3; birds have 2-3-4-x-x ("x" means that even the metacarpal is missing) to 1-2-1-x-x. Pterosaurs have 2-3-4-4-x; the reason it isn't 2-3-4-5-x is that the claw of the wing finger, along with the bone that supported it, is lost.
Your explanation, on the other hand, cries for an explanation why the middle 3 fingers each lost one phalanx (something that doesn't normally happen) while the 5th gained one (something that... has never happened in amniotes except hadrosauriforms, I think).
And?
So straightforward that scientists thought about it in the late 19th century and then dismissed it summarily because it is too simple -- it fails to account for... for the existence of any dinosaurs other than birds, for instance!
<sigh>
That you don't know them doesn't mean they don't exist, and it doesn't even mean that other people don't know about them.
Please!
Then go to pterosaur.net as recommended in comment 133, and read books on vertebrate anatomy and vertebrate paleontology -- not just the tiny, tiny summaries on Wikipedia!
Jack:
"Some pterosaurs had 4-toed feet, like birds."
David MarjanoviÄ:
"And?"
And that is like birds. Which is what we are talking about.
Also people are misunderstanding what I said about the fusing of bones.
I am not saying they were fused in pterosaurs. I am saying they fused in the transition from pterosaur to (basal) bird.
Jack:
"The pteroid bone is the evolved form of the thumb."
David MarjanoviÄ"
"What makes you think so?
Check out the way it looks, and the place where it articulates with the wrist."
I have looked.
As I said earlier:
How can we definitively decide this question? I am not much interested in just trading opinions.
Can we definitively decide this question?
And if it is acknowledged that we cannot definitively decide this question, then let that be acknowledged.
David MarjanoviÄ:
"Well... no. They're not fused in any pterosaurs, or for that matter in any birds!"
http://en.wikipedia.org/wiki/Bird_anatomy
"The shoulder consists of the scapula (shoulder blade), coracoid (see The Chest), and humerus (upper arm). The humerus joins the radius and ulna (forearm) to form the elbow. The carpus and metacarpus form the "wrist" and "hand" of the bird, and the digits (fingers) are fused together. The bones in the wing are extremely light so that the bird can fly more easily."
Some of the fingers are fused in birds.
Also note: The thumb - the alula - is still there of course. It is called the "pteroid bone" in pterosaurs.
I am sure everyone here knows about the alula. It is such a marvelous piece of engineering. Using the ancestral thumb (evolved from the pterosaur) to add fine control to wing movement.
http://en.wikipedia.org/wiki/Alula
"The alula, or bastard wing, is a small projection on the anterior edge of the wing of modern (and some ancient) birds. The alula is the freely moving first digit, a bird's "thumb," and is typically covered with three to five small feathers, with the exact number depending on the species".
David MarjanoviÄ:
"The normal phalangeal formula for amniote hands is 2-3-4-5-3 (2 phalanges in the thumb, 3 in the index finger, and so on). Dinosaurs have at most 2-3-4-3-3; birds have 2-3-4-x-x ("x" means that even the metacarpal is missing) to 1-2-1-x-x. Pterosaurs have 2-3-4-4-x; the reason it isn't 2-3-4-5-x is that the claw of the wing finger, along with the bone that supported it, is lost."
I am presenting the idea that the pterosaur had five fingers. The pteroid bone is the thumb. The difficulty with any listing of the phalanges such as David has given is that it ignores the pteroid bone which is the thumb, and starts counting from the index finger as if the index finger were the thumb.
David MarjanoviÄ:
"Better yet, the finger that supports more than half of a pterosaur wing is the 4th, and that finger is completely missing in birds."
Here again we see the confusion that occurs when the pteroid bone (the thumb) is ignored. For example, it is not the 4th finger that supports more than half of a pterosaur wing. It is the 5th.
When comparing fingers and their phalanges, the pteroid bone must be included as the thumb. (And of course it must be kept in mind that in birds some of the fingers are fused).
Well done Jack. You have successfully turned the comments into idiot corner. You clearly do not know the first clue about the subject. Even if the pterosaur pteroid is the same as the thumb (and is almost certainly is not) do you really think that this overturns the theropod origin of birds? Hundreds of features are involved, not two or three.
Thanks David, for taking the time to explain stuff like the finger formula! I'm surprised time and again at the kinds of things I can learn here. :)
Simon Spaulding:
"Well done Jack. You have successfully turned the comments into idiot corner. You clearly do not know the first clue about the subject. Even if the pterosaur pteroid is the same as the thumb (and is almost certainly is not) do you really think that this overturns the theropod origin of birds? Hundreds of features are involved, not two or three."
Simon, you say there are "hundreds of features".
Could you list the ten that you take to be the most important? Or if you cannot evaluate which are most important, please list ten or twenty (of the hundreds). I would be delighted to address them.
I have addressed the bulk of the issues that have been mentioned so far.
Also concerning whether the pteroid bone is the thumb - how can we definitively decide this question? Can we definitively decide this question?
And if it is acknowledged that we cannot definitively decide this question, then let that be acknowledged.
I want to approach this in an evidence-based way and not just trading opinions.
Jack: Please explain just how the many similarities between theropods such as Velociraptor or even Compsognathus are in any way less convincing than any perceived similarity between pterosaurs and birds.
I note that he's not answer my question, which is very similar to yours, Hai~Ren.
Hello Hai~Ren.
I am addressing what people believe are the differences between pterosaurs and birds. Many people believe mistakenly that there are many significant differences.
I am not wishing to be sidetracked into a discussion about the theropod alternative. That can be done later.
Are there any pterosaur to bird differences that are significant to you?
There have been references to the ilium, pubis and ischium.
Birds and pterosaurs are very similar in this area:
http://icb.oxfordjournals.org/cgi/content/full/40/1/87
"Several aspects of the pelvic girdle suggest that pterosaurs were specialized for pelvic aspiration during flight. As in birds, the three pelvic bones of pterosaurs were solidly fused into a single unit (Fig. 9), and an increased number of dorsal vertebrae were incorporated into the sacrum (3 to 5 in Rhamphorhynchus and as many as 10 in Pteranodon) (Wellnhofer, 1978Go, 1987Go). The preacetabular process of the ilium was very long and in pterodactyloids it was fused with additional dorsal vertebrae to form a synsacrum-like structure.
Look, Jack, the point is that theropods already HAVE a bunch of birdy features, moreso than in pterosaurs. Why posit pterosaurs as an ancestor when theropods are already a better alternative? It's like going backwards.
Also, have you seen a pterosaur pelvis? Have you seen Archaeopteryx's pelvis? Have you seen a dromaeosaur pelvis? You tell me which ones look the most similar. Also, you're forgetting that pterosaurs have a prepubis bone, something unique to the group, somethings birds do NOT have.
Also, birds have an open acetabulum whereas pterosaurs do not. This may be problematic to your idea.
I may not be a paleontologist, but I do know that that is not what "parsimonious" means.
The sesamoid in panda bears is not an "evolved thumb" (in the sense of being derived from a digit of the hand), and it is not "parsimonious" to assert that it is.
Study its anatomy a little more carefully than just glancing at a single line in a single paragraph in a single paper.
Do modern birds have an open acetabulum?
If so, please provide references.
Here is all I could see on this:
http://en.allexperts.com/q/Paleontology-Dinosaurs-1571/modern-birds-sti…
"The perforate acetabulum in dinosaurs is shared by primitive birds, including Archaeopteryx, but the feature is not present in modern birds. In fact the entire pelvis of birds has undergone extensive evolution since the days of Archaeopteryx, with the ilium, ischium and pubis having fused into one piece (the innominate bone). The hole which was once present closed up prior to the appearance of the group to which all modern birds belong (the Neornithes).".
Owlmirror.
I am not talking about pandas.
I have asked:
"Is there some way to definitively disprove that idea?"
Can you answer that question? I have asked it a few times and nobody has answered it.
I am wanting to discuss this based on facts and evidence - not opinion.
I know. It was an analogy. Do you know what an analogy is?
All that you've offered has been your uninformed opinion.
Here is what I said earlier:
"Also people are misunderstanding what I said about the fusing of bones.
I am not saying they were fused in pterosaurs. I am saying they fused in the transition from pterosaur to (basal) bird."
I see that this can be misunderstood.
I am not saying that pterosaurs evolved into "basal" birds as they are defined by the dino to bird view.
I am saying pteroaurs evolved into the earliest modern birds.
Right. Because you don't actually know what "parsimony" means.
Or rather, you've been offering an incoherent argument from deep ignorance about pterosaurs and birds and evolution.
Do you know the difference between analogous and homologous structures? Please present facts and evidence that demonstrate that you have this knowledge -- not just your opinion.
David MarjanoviÄ:
"The normal phalangeal formula for amniote hands is 2-3-4-5-3 (2 phalanges in the thumb, 3 in the index finger, and so on). Dinosaurs have at most 2-3-4-3-3; birds have 2-3-4-x-x ("x" means that even the metacarpal is missing) to 1-2-1-x-x. Pterosaurs have 2-3-4-4-x; the reason it isn't 2-3-4-5-x is that the claw of the wing finger, along with the bone that supported it, is lost."
I have a question.
In saying that birds have 2-3-4-x-x is that taking the alula as the thumb? In other words does the alula have the 2 phalanges? Can you provide a reference (a link) for this.
Jack, are you suggesting that basal birds ARE dinosaurs, but modern birds are pterosaurs? You're suggesting that birds evolved TWICE?
Basal birds are not dinosaurs. Birds did not evolve twice.
Modern birds evolved from pterosaurs.
Um... do you know what the term "basal" means in this context?
Concerning pterosaur clavicles:
http://books.google.com/books?id=8CKYxcylOycC&pg=PA206&lpg=PA206&dq=pte…
"In pterosaurs the clavicle was incorporated into the sternum."
"David Unwin, 2006. The Pterosaurs from Deep Time."
"Although it looks like a single bone, studies of an immature pterosaur show that it was made up of several elements, including the clavicles, which were thought to have been lost, but now appear to have been hiding in the breast bone all along."
He's referring to Wild, R. 1993. A juvenile specimen of Eudimorphodon ranzii Zambelli (Reptilia, Pterosauria) from the Upper Triassic (Norian) of Bergamo. Rivista del Museo civico di Scienze Naturali âE. Caffiâ Bergamo 16:95-120
(I do not have a link for this).
"Basal birds are not dinosaurs. Birds did not evolve twice.
Modern birds evolved from pterosaurs."
All hope is lost.
For an encore, he will no doubt conclude that bats evolved from bugs.
References:
Calvin and Hobbes. 1989. Bats: The Big Bug Scourge of the Skies. Communications from Elementary School Studies. 1989-11.
Jack, what you need is the <blockquote> tag: this
<blockquote>quoted text here</blockquote>
is displayed this way:
Sure it is, but it's also like ornithopods (hadrosaurs excluded), like Keraterpeton the Carboniferous "nectridean" amphibian, like hippos, like pigs, like... early ichthyosaurs, like... crocodiles... I could go on for an hour.
What do you mean by "definitively"?
In case you mean absolute certainty of the kind found in mathematics, nothing in science can be decided definitively.
In a question of how evolution proceeded, all we can make are arguments from parsimony: those hypotheses that require the smallest number of additional assumptions must be preferred.
Wikipedia is wrong about this.
OK, you might argue that the 3rd finger is fused to the 2nd in modern birds, because it's so short that it doesn't stick out anymore; but even there, the bones aren't fused.
Why do you think that the pteroid is the thumb? Why?
I've presented two arguments against that interpretation: it's not in the right position, and the phalangeal formula fits better otherwise. You have not addressed this.
This is ridiculous, because ideas about how evolution has proceeded can only be evaluated by comparing them to each other -- by finding out which one is more parsimonious than all others.
They simply can't be judged on their own. How would one do that?
Yes!
That the alula is the thumb is textbook wisdom. It's not new, and hasn't been for well over 100 years.
Yes! But you omitted the part I wrote behind the parenthesis: 2-3-4-x-x is the maximum for birds (found, for instance, in Archaeopteryx and Confuciusornis); modern flying birds can have as little as 1-2-1-x-x. That is, the 2nd bone of the alula (with the claw that it often supports) is sometimes lost. Sometimes, but not always; check out the next few chicken and turkey wings that land on your dinner table! Many have a claw on the 1st and/or 2nd finger.
Fine. Why would this fusion ever reverse? And why do we need such a contrived explanation when all theropods (where it's preserved) have a wishbone, and Massospondylus has clavicles that touch in the middle?
Okay, compare the idea I have presented concerning the pteroid bone as the thumb, with your favorite alternative and show us that your alternative is more parsimonious.
If you do not want to do the comparison, then just describe the alternative that you believe is more parsimonious.
In the development of the pterosaur the clavicles become part of the breastbone. All that is required is that it not become fused to the breastbone in the early part of the development process.
So here is the comparison of finger phalanges of pterosaur to bird when the pteroid bone is considered the thumb.
Pterosaur: 1-2-3-4-4.
Modern Bird 1-2-1-x-x
Would you agree?
And I take it that there are variations in these numbers as well in different species.
Jack - since April 14th you've posted 30 comments here. You show no indication of understanding the counter-arguments to your proposal, or of giving up on an idea that is clearly not in the least bit parsimonious. Please stop now - I am closing comments on this article. Please take away the following points...
-- If organisms (like pterodactyloid pterosaurs and birds) share anatomical characters, that might be because those characters are widespread within ornithodirans, within archosaurs, or even within diapsids or reptiles or amniotes. You need to understand the difference between 'primitive' and 'derived' characters.
-- If organisms (like pterodactyloid pterosaurs and birds) share superficially similar characters (like toothless rostra), that might be because those two groups have convergently evolved similar solutions to similar problems. In this case, comparison of early forms with more advanced ones shows that convergence is indeed the correct interpretation (viz, advanced birds and advanced pterosaurs both share toothless rostra, but primitive birds and primitive pterosaurs have teeth).
-- Because animals are complicated objects that are composed of hundreds of anatomical components, it is not satisfactory to use one or two superficially similar characters to imply a relationship: you need to analyse tens or hundreds (or more) of such features. This is now routine among people who study phylogeny.
-- A substantial literature shows that birds are maniraptoran theropod dinosaurs: this is because birds share numerous derived anatomical characters with other theropods. In turn, theropods are saurischians, and saurischians are dinosaurs.
-- You cannot claim on the one hand to have read the literature, and then say that you are not aware of the literature concerned. At the very least, you need to be familiar with Gauthier (1986), Holtz (2000) and Ostrom (1976). Pterosaurs do not have a close relationship with birds: birds and pterosaurs do not share any derived characters not seen in other ornithodirans, and pterosaurs lack the characters that might place them within Dinosauria, Saurischia, Theropoda or Maniraptora.
Refs - -
Gauthier, J. 1986. Saurischian monophyly and the origin of birds. Memoirs of the California Academy of Science 8, 1-55.
Holtz, T. R. 2000. A new phylogeny of the carnivorous dinosaurs. Gaia 15, 5-61.
Ostrom, J. H. 1976. Archaeopteryx and the origin of birds. Biological Journal of the Linnean Society 8, 91-182.