
tags: researchblogging.org, endangered species, estimating extinction risk, demographic heterogeneity, demographic stochasticity, environmental stochasticity, Mechanistic stochastic models, Brett Melbourne

The endangered pelagic thresher shark, Alopias pelagicus.
More than half of the world's shark species are under the threat of extinction
due to overfishing by humans, especially for sharkfin soup.
Image: Kevin Markey, 2004 (Pacific Shark Research Center).
![]()
The International Union for Conservation of Nature (IUCN) released a report in 2007 indicating that more than 16,000 animal species worldwide are threatened with extinction. These numbers translate into one in four mammal species, one in eight bird species and one in three amphibian species that are included on the IUCN's "Red List" of endangered species. However, a paper was just published suggesting that the risk of extinction for many animals and plants has been underestimated by as much as 100-fold. According to the paper's lead author, assistant professor Brett Melbourne of University of Colorado at Boulder's ecology and evolutionary biology department, this underestimation was the result of a mathematical "misdiagnosis."
The extinction risk model currently in use is based primarily on two factors: the number of random events adversely affecting individuals within a population, such as an accidental death, and the impact of random environmental events such as temperature and rainfall fluctuations that can influence birth and death rates.
But the current model ignores random differences between individuals within a given population at a given place and time. These differences include variations in gender ratios (sex), size variations within a species, and behavioral differences (demographic heterogeneity), randomness in births and deaths and reproductive success (demographic stochasticity), and random environmental effects upon the birth-death rate (environmental stochasticity) (Figure 1);
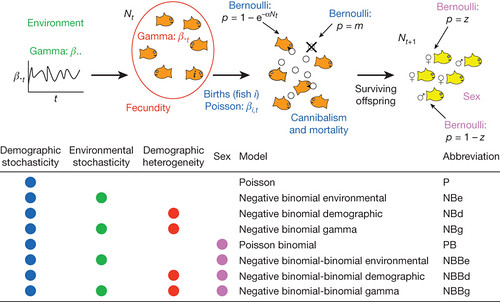
Figure 1: A family of stochastic Ricker models based on Ricker's26 assumptions about the life cycle of a fish species that cannibalises its eggs.
The stochastic models incorporate stochasticity in various parts of the life cycle, including gamma variation in environmentally determined birth rates, gamma variation in birth rates between individuals, Poisson variation in birth rates within individuals, Bernoulli variation in mortality within individuals, and Bernoulli variation in the sex of an individual at birth. [larger view].
"There has been a tendency to misdiagnose randomness between individuals in a population by lumping it with random factors in the environment, and this underestimates the extinction threat," reports Melbourne. This is much like miscalculating the odds in an unfamiliar game of cards because you didn't know the rules, he explained.
Melbourne and his colleague, Alan Hastings, 2006 MacArthur Award winner and professor in the department of environmental science and policy at the University of California, Davis, developed a more inclusive mechanistic stochastic model for estimating extinction risk that analyzes all these previously ignored differences.
To develop and test their mathematical model, Melbourne and his colleague monitored populations of red flour beetles, Tribolium castaneum, in lab cages and used the results to test their new mathematical models (Figure 2);
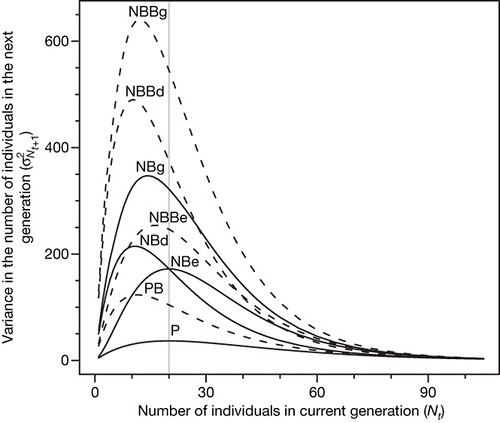
Figure 2: Variance in the number of individuals in the next generation (Nt + 1) as a function of the number of individuals in the current generation (Nt) for the stochastic Ricker models.
The model parameters were: R = 5, α= 0.05, kD = 0.5, kE = 10. The stochastic parameters (kD, kE) were set so that the total variance due to demographic heterogeneity was equal to the total variance due to environmental stochasticity. The vertical bar indicates the position of the stationary point in the Ricker production function. Abbreviations identify the models listed in Fig. 1. [larger view].
Some of these stochastic factors, such as the beetles' sizes and the sex ratio of the population, could explain large swings in a species population that were unaccounted for by the older model. Their full stochastic model was sensitive to the relative importance of environmental and demographic variability, and therefore it accurately predict extinction risk for their beetle populations (Figure 3);
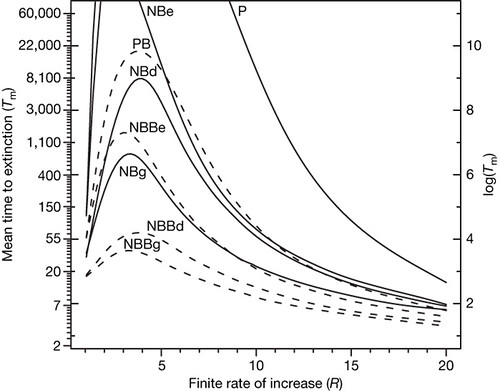
Figure 3: Intrinsic mean time to extinction 30 (Tm) for the stochastic Ricker models as a function of the finite rate of increase (R).
The model parameters were: kD = 0.5; kE was adjusted so that the total variance in Nt + 1 due to demographic heterogeneity was equal to the total variance due to environmental stochasticity; α was adjusted to hold the equilibrium density at 30 individuals. Internal ticks show the natural logarithm scale. Abbreviations identify the models listed in Fig. 1. [larger view].
Natural animal populations, even when they are very small, are more likely to have larger differences in sex ratios and differences between individuals than laboratory-controlled beetle experiments. As a result, "the effect we have uncovered here will be larger in natural populations," the authors point out in their paper.
This new model will yield immediate benefits. For example, even though biologists can collect data on specific individuals to help develop and track extinction trajectories for large, high-profile terrestrial species like Sumatran tigers, marine fishes and mammals are a different matter.
"[F]or many other species, like stocks of marine fish, the best biologists can do is to measure abundances and population fluctuations, and it's these species that are most likely to be misdiagnosed," observes Melbourne.
"When we apply our new mathematical model to species extinction rates, it shows that things are worse than we thought," said Melbourne. "By accounting for random differences between individuals, extinction rates for endangered species can be orders of magnitude higher than conservation biologists have believed."
Source
Melbourne, B.A., Hastings, A. (2008). Extinction risk depends strongly on factors contributing to stochasticity. Nature, 454(7200), 100-103. DOI: 10.1038/nature06922.
University of Colorado at Boulder press release (quotes).
Yay! Someone else uses fish when talking about Poisson processes.
I printed out the paper yesterday, and read it over lunch. Unfortunately all the technical action was in the supplementary information. I guess that will wait until lunchtime today.
It's cool stuff, though.
i'd be most interested to read your comments after you've read the supplementary information.
OK, here you go...
The basic idea of incorporating inter-individual variation is an important one, and that's why I found it exciting. There are some technical issues I have. I guess the most important one is also the most technical - I'm not sure they are calculating the total variance appropriately, which means that their figures (the two graphs above) might not be quite right. I don't think this changes the overall message, but the ordering of the models might change.
I'm not sure how much of a problem this is in practical conservation, because I don't know enough about the models they use. There are some models where it shouldn't be a problem, because the demographic heterogeneity will be estimated as demographic stochasticity, and there's no problem. I don't know how often they are used, though. Perhaps I should ask.
There's a lot more that could be done, based on this paper - it would be the natural extension of some of the things I've been doing. It would be nice to see how things go in the field - they suggest that demographic heterogeneity will have a stronger effect compared to the lab, but so will environmental stochasticity.
But most importantly, the names of their distributions are nothing as compared to my zero-truncated, one inflated negative binomial distribution.