
tags: researchblogging.org, evolution, speciation, ring species, phylogeography, landscape genetics, crimson rosella, Platycercus elegans, parrots, birds, Australia

![]()
One of the challenges facing those who believe that evolution cannot create new species is explaining the problem of "ring species." Ring species are a group of geographically connected populations that can interbreed with nearby populations, but cannot breed with those populations that exist at each end of the cline (figure A). These populations are known as "ring species" because their populations often form a ring where each end of the cline is located near the other (figure B), or the ends of their ranges actually overlap (figure C), thereby curling inwards upon each other and forming a ring.
Ring species also provide an interesting conundrum to those who would divide the world into discrete species since all that distinguishes a ring species from two separate species is the existence of connecting populations -- but if enough of the connecting populations within the ring perish, gene flow is severed, and the ring species are then isolated into two distinct species.
Currently, the Greenish Warbler species complex, Phylloscopus trochiloides, that forms a ring around the Himalaya Mountains is the best documented example of a ring species complex. Several other species complexes that were once thought to be ring species are the Larus gulls that live along the coastlines of the Pacific Ocean, and another Larus group that live along the shores of the Atlantic Ocean. Less familiar to bird watchers who live in Europe and North America is another proposed ring species complex consisting of parrots: the Crimson Rosella, Platycercus elegans species complex of Australia.
The Crimson rosella, Platycercus elegans, is easy to identify because of its almost surreal crimson and blue plumage. But closer examination shows that there are different color schemes across this bird's range throughout southern and eastern Australia. In fact, in some areas, red is replaced with bright yellow -- a form known as the Yellow Rosella -- while in other populations, some of the red on the back and cheeks is replaced with a fiery yellow-orange -- the Orange Rosella.
"The yellow birds are found in the drier, inland areas, so the color of their plumage may have evolved to blend in with their habitat," observes reports Gaynor Dolman of Australia's Commonwealth Scientific and Industrial Research Organisation's (CSIRO) National Wildlife Collection and co-author of research conducted by an international team that was just published in the prestigious journal, Proceedings of the Royal Society B.
''We don't know, but the redder birds might have evolved those colors so they could pick out their mates among the greener vegetation," she said. "Or, the colors could have something to do with what female birds find attractive.''
However, so striking are the different colors and gradations between them within this group of parrots that research from the 1950s suggested that the Crimson Rosellas are an example of ring species. For example, the population forms a circle around the less habitable semi-arid regions of Australia where they don't live, with the two ends of the ring joining near the headwaters of the Murray and Murrumbidgee rivers in the Snowy Mountains.
"Many evolutionary biologists have argued that the different forms of Crimson Rosellas arose, or speciated, through 'ring speciation'," reports Dolman.
To determine if the Crimson Rosella species complex is a ring species, the team analyzed mitochondrial (mtDNA) and chromosomal, or nuclear, DNA (nDNA) from blood and tissue samples collected from 307 unrelated adult rosellas from 92 localities (figure 1);

Figure 1. (a) Distribution and plumage phenotype variation in parrots of the Crimson Rosella complex in eastern Australia (from Forshaw & Cooper (2002) in this and subsequent figures). Note the narrow zone of unsuitable habitat currently separating Adelaide and Yellow. (b, c) Sampling scheme of vouchered museum specimens for mtDNA analyses of the full mainland distribution of the complex. [larger view].
DOI: 10.1098/rspb.2008.0765.
The team's DNA analyses revealed that the Crimson Rosella species complex is monophyletic in origin -- essential for ring species, and the populations located north of the Hunter River form a distinct group from those birds that occur in the south and southeastern regions of Australia. However, the DNA analyses combined with the birds' geographic ranges revealed a much more complicated picture than the research team anticipated (figure 2);

Figure 2. Unrooted network of mtDNA diversity in southeastern Australian populations of the Crimson Rosella complex, showing groups (a) 1, (b) 2 and (c) 3 as referred to in the text. Open circles indicate unsampled haplotypes. Colours in circles indicate the plumage phenotype of samples as in figure 1. The sample size for each haplotype is 1, unless otherwise indicated with a number. KI refers to haplotypes found in eight Kangaroo Island Crimson individuals. A solid line with open circles shows where the North Queensland populations join the network. A unimodal mismatch plot for (c) is shown in (d). (d) Dotted line, observation; solid line, expected. [larger view].
DOI: 10.1098/rspb.2008.0765.
For example, the team's analysis showed that Crimson Rosellas from Adelaide (which are either crimson or orange in color) and those from the Western Slopes near Wagga Wagga (also crimson or orange) had inexplicable genetic similarities (figure 2b, c) even though they are located 200 kilometers apart. Additionally, Crimson Rosellas on Kangaroo island were genetically similar to the Adelaide Rosellas, despite these two populations being separated by 15 kilometers of ocean -- a geographical barrier that significantly slows gene flow. Curiously, the geographically linked Yellow and Orange Rosella populations in the inland areas of southeastern Australia showed distinct genetic differences -- differences that should not be present if they are part of a ring species complex.
The team then focused their efforts on just the southeastern populations of the Crimson Rosellas, which might qualify as a ring species. They examined their microsatellite genotypes to identify patterns in gene flow and plumage coloration (figure 3);
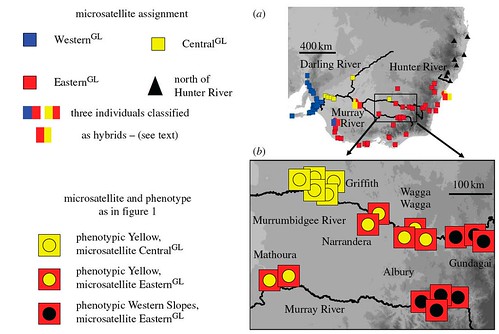
Figure 3. (a) Summary of microsatellite analyses of southeastern Australian populations of the Crimson Rosella complex. Coloured squares are microsatellite-based GENELAND classifications of individuals as described in the text (EasternGL, CentralGL and WesternGL). Bicoloured symbols show the three individuals that were classified as 'hybrid' assignments of 0.61 EasternGL/0.38 CentralGL, 0.05 EasternGL/0.95 CentralGL and 0.16 EasternGL/0.84 WesternGL. (b) The map details sampling for microsatellites in the upper and middle reaches of the Murray River system and shows discordance between microsatellitebased assignments (squares) and plumage phenotypes (circles). [larger view].
DOI: 10.1098/rspb.2008.0765.
Unfortunately, the data did not reveal any conclusive relationships between DNA, plumage color and geographic location for these populations, contrary to the prediction by the ring species hypothesis.
"We found that in the case of Crimson Rosellas, their three separate genetic groups don't show a simple link to the geographical distribution of the color forms," reports Dolman.
Their research identified three distinct genetic clusters among this population -- contrary to what they should find if crimson rosellas are ring species. Oddly, the first genetic discontinuity was identified within the Yellow Rosella population rather than further east, where there is a visible color transition between the Crimson and the Yellow Rosellas. Further, this genetic discontinuity occured in the absence of any geographical barriers to gene flow.
The second genetic discontinuity was identified farther west, between the Adelaide and Yellow rosella populations. This is consistent with current geographical discontinuity but was not predicted by the ring hypothesis.
The third genetic discontinuity that was not predicted by the ring species hypothesis is perhaps the oddest of all. This genetic discontinuity was identified between the Adelaide and Kangaroo Island Crimson rosella population on the one hand and mainland Crimson rosella population on the other. The team's data did not detect the 15 kilometer sea gap between the Adelaide and Kangaroo Island Crimson rosellas as a genetic discontinuity (figure 3), while the data did identify the 200 kilometers of unsuitable habitat along the mainland coast that isolates Adelaide from mainland Crimson (figure 1).
''[W]e had to reject that [ring species] theory because we found multiple genetic discontinuities around the ring. We found the three separate genetic groups didn't show a simple link to the geographical distribution of the three different color forms,'' Dolman reports.
Despite the fact that the data clearly identified three genetic discontinuities in the southeastern Crimson Rosella species complex that are inconsistent with predictions made by the ring species hypothesis, the researchers cannot -- yet -- completely rule out that Crimson Rosellas did not evolve as a ring species.
''It is possible the Crimson rosellas formed a ring at some stage in their evolutionary history, but the evidence could have been lost through changes to the climate or environment,'' Dolman concedes.
For example, Australia's Crimson Rosellas could have evolved their different plumage colors due to geographic barriers presented by rising sea levels and the disappearance of an ancient, vast freshwater lake in the Murray River basin of Australia.
''It shows we still have a lot to learn about some of most our most familiar species that we take for granted. There's still a lot of evolutionary puzzles to solve,'' Dolman concluded.
Sources
Joseph, L., Dolman, G., Donnellan, S., Saint, K.M., Berg, M.L., Bennett, A.T. (2008). Where and when does a ring start and end? Testing the ring-species hypothesis in a species complex of Australian parrots. Proceedings of the Royal Society B: Biological Sciences, -1(-1), -1--1. DOI: 10.1098/rspb.2008.0765.
CSIRO press release (quotes).
The Larus ring complex is now not thought to be a ring species either:
Liebers, Dorit, Peter de Knijff, and Andreas J. Helbig. 2004. The herring gull complex is not a ring species Proc. R. Soc. Lond. B 271: 893-901.
Now that is one gorgeous bird. The richness of the coloring is quite stunning.
I'd think that what humans perceive as a ring may differ from what a bird might consider as a ring, and I doubt we will ever find any species which fits exactly the concept of ring species, but it's useful in the general sense.
Enjoyed your article. Very interesting. I am a long-time bird keeper who has owned Rosellas for 20 years. I have had several different species and their interest in each other has to do more with their individual personalities (e.g., bossy, less bossy -- these are bossy birds) than whether a particular bird is of the same species. There are many mutations and hybrids available among Rosellas, as well. My three eastern rosellas differ significantly in feather density, size, and in how well defined their markings appear.