
tags: neurobiology, neuroscience, animal communication, birdsong, premotor nucleus HVC, brain temperature, neural circuitry, motor behaviors, bioacoustics

Captive-bred Zebra Finch, Taeniopygia guttata, at
Bodelwyddan Castle Aviary, Denbighshire, Wales.
Image: Adrian Pingstone/Wikipedia [larger view].
Birdsong is the primary model system that helps scientists understand how the brain produces complex sequences of learned behavior, such as playing the piano. In songbirds, there are many interconnected brain regions that play specific and important role in the production of song. These brain regions are known as "song control nuclei." Two song control nuclei, the High Vocal Center (HVC) and the robust nucleus of the arcopallium (RA), are especially important for birdsong because damage to either prevents the bird from singing altogether. It was hypothesized that either HVC or RA could orchestrate the rhythm of the bird's song, acting like a sort of biological metronome, but the technology did not exist to identify which song control nucleus might play this role -- until now. In an elegant combination of physics and biology, two researchers at Massachusetts Institute of Technology in Cambridge invented a tiny new device that allowed them to identify HVC as regulating the rhythm of birdsong.
Neurons send messages more slowly when chilled, so the researchers hypothesized that, by cooling only specific song control nuclei, they could selectively slow the firing rate of each region in turn to slow function.
"Temperature has a very strong effect on the speed at which neurons operate," explains Fee. "We realized we could use temperature to very subtly control the speed of the circuit and see whether that affected the timing of singing."
To do this work, Michael Long and Michale Fee invented a tiny Peltier cooling device that is similar to portable electronic beverage coolers (figure 1a). Then they used this device to produce a small temporary cooling effect that could be localized to precise parts of the brain (figure 1acb);
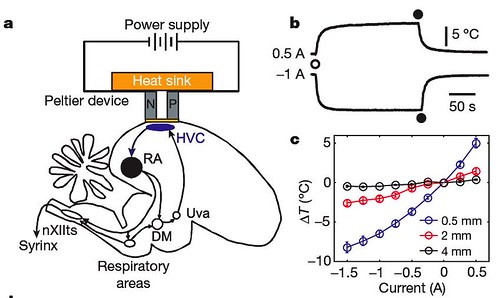
Figure 1 | Changes in HVC temperature affect song duration. a, The Peltier device and relevant parts of the song production pathway. N and P, semiconductor elements; DM, dorsomedial nucleus of the intercollicular complex; nXIIts, tracheosyringeal part of the hypoglossal nucleus. b, Temperature change in HVC as a function of time after onset (open circle) of the indicated current through the Peltier device (top, heating; bottom, cooling); current switched off at filled circles. c, Calibration curves for brain temperature changes (ΔT) at various depths under the Peltier device (n54) [larger view].
DOI: 10.1038/nature07448.
"We suspected that cooling different brain regions involved in singing might alter the song in different ways," explained Long, a postdoctoral researcher in the Fee lab.
Male zebra finches, Taeniopygia guttata, produce a short "beeping" song that lasts roughly one second. When Long and Fee recorded the bird's song after a targeted brain region had been cooled, and compared that song to the bird's song when that brain region was at normal body temperature, they found that chilling HVC produced a dramatic change in the bird's song: slowing the song without changing the overall quality of the individual notes produced (figure 1d);
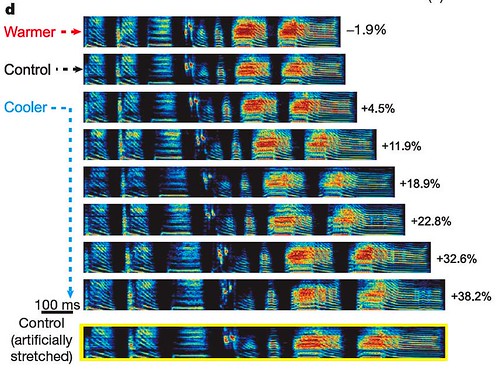
Figure 1 | Changes in HVC temperature affect song duration. d, Representative sonograms (frequency, 1-9 kHz) recorded from bird no. 3 with HVC heated (0.25 A) and cooled (0.25 to 1.5 A in 0.25-A steps), showing percentage song dilation relative to control. Hotter colours represent greater sound intensity. Bottom, spectrogram of the control motif shown artificially stretched. [larger view].
DOI: 10.1038/nature07448.
You can hear how these sonograms sound when played, and you can visually and auditorially compare the experimental birds' songs (below);
|
Control Song (normal body temperature) Stretched Song (HVC cooled by 8.2° C) |
"We ran the current through the cooling device, put a female in front of a male, he started singing and it came out more slowly," recalls Fee. "It was amazing. The effect was very clear."
As you can see and hear from the above data, the overall rhythmic structure of the bird's song was preserved without changing the notes within the song. In fact, the only change was the overall increase of the song's duration. This effect was consistent across all experimental birds (figure 1ef);
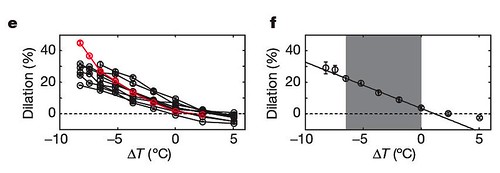
Figure 1 | Changes in HVC temperature affect song duration. e, Percentage change in duration (dilation) of song motif versus change in temperature, relative to the pre-implantation song (n510). Red, bird no. 3. f, Motif dilation averaged over all ten birds. The shaded area represents the range over which the song stretch metric (slope) was calculated. All error bars, s.e.m. [larger view].
DOI: 10.1038/nature07448.
These data show that the bird's song slowed in proportion to the degree of cooling, with the biggest temperature change (a 10 degrees Celsius reduction) causing the song to stretch out by roughly 30 percent.
But these dramatic effects were only seen when HVC was cooled: cooling RA had almost no effect on the bird's song (figure 3);
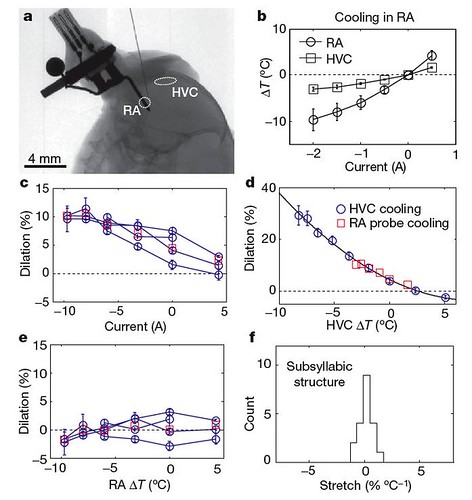
Figure 3 | Effects of RA temperature change on song timing. a, X-ray image
of the implanted RA cooling device and approximate locations of HVC and RA. b, Temperature in RA (200 mm from cooling probe) and HVC as a function of RA probe current. We note that the RA probe produces some cooling in HVC. c, Change in motif duration as a function of RA probe current (n54; red squares, mean). d, Average change in motif duration (red squares) during RA probe cooling or heating, plotted as a function of HVC temperature. Also plotted is the average change in motif duration (blue circles) as a function of HVC temperature measured in the HVC cooling experiment (Fig. 1f). e, Change in motif duration as a function of RA temperature, corrected for the effect of HVC temperature change. All error bars, s.e.m. f, Stretch of subsyllabic elements for the population of RA cooled birds (n54, 20 syllables), corrected for HVC temperature change [larger view].
DOI: 10.1038/nature07448.
"It has been very difficult to figure out where in these different parts of the brain the timing is generated," says Fee. "It wasn't clear that cooling any of these brain areas wouldn't just make the circuit break. It was surprising to find that when we cool the HVC the song just stretches out."
HVC is a bilateral structure, with these region near the brain surface on each side of the forebrain, so Fee and Long wondered how cooling only one HVC might affect timing of the birds' songs: does one HVC function as the "master clock" while the other is the "slave"? Or does one HVC control timing of some parts of the song, while the other HVC controls other song elements?
Long and Fee's data (not shown) revealed that that chilling only one HVC resulted in an intermediate slowing of the birds' songs, and further, they found that some song elements that were not stretched by right HVC cooling were strongly stretched by left HVC cooling, and vice versa. So based on these data, Long and Fee concluded that each HVC regulates the timing of certain portions of birdsong.
Fee and his colleagues are now using this cooling device to elucidate precisely how HVC controls song timing. Their previous electrical recordings of individual HVC neurons suggest it functions like a cascade of falling dominoes, with waves of activity propagating at a fixed speed through the neural circuitry -- an idea they are now testing.
"It seems to look a lot like a music box," Fee says. "You have a drum that rotates and controls the timing. The drum has little bumps all over it. If you rotate the drum more slowly, you get the same notes but they come out more slowly."
This innovative new cooling technology can also be used in other animals to elucidate a variety of complex behaviors.
"We can also use this cooling technology to discover which brain regions control the timing of different complex behaviors in different animals, something that has been very difficult to assess until now," Fee said. "We know that HVC is related in some ways to [the] human cortex, so it could be showing us a very general mechanism for representing the passage of time within the brain."
In short, scientists have almost no detailed information about how speech, language and musical performance are encoded as motor commands. But by studying songbirds, we can getting a small glimpse of how the general mechanisms for vocal learning operate.
Sources
Michael A. Long, Michale S. Fee (2008). Using temperature to analyse temporal dynamics in the songbird motor pathway Nature, 456 (7219), 189-194 DOI: 10.1038/nature07448.
MIT press release (quotes).
Thanks for a great overview of some fascinating research. I do have one question, though...
they found that chilling HVC produced a dramatic change in the bird's song: slowing the song without changing the overall quality of the individual notes produced
What do you mean by the "overall quality" of individual notes? It looks from the spectrogram that the notes have qualitatively changed, i.e. the notes are elongated. Slowing the song could mean a number of different things. I could play "Happy Birthday" more slowly by:
1) Increasing the lengths of the intervals between notes
2) Increasing the lengths of the notes, but keeping the intervals constant
3) Increasing both the lengths of the notes and the intervals
It looks like the cooling manipulation to HVC resulted in #3 in the birds, which is interesting. I don't think it's intuitive, either. Do the authors elaborate on this?