
The question of how birds migrate long distances has long baffled researchers, and there are various hypotheses about which navigational cues birds use when migrating. Over the years, it has been suggested that migrating birds use smell, visual cues such as the position of the sun, the geomagnetic field, or a combination of these. It is, for example, known that induced magnetic fields and electrical storms disrupt the navigational abilities of homing pigeons, but exactly how the birds detect, perceive and interpret a magnetic field remains a mystery.
A recent study led by Todd Dennis, of the Department of Biological Sciences at the University of Auckland, New Zealand, provides compelling evidence that the Earth's magnetic field alone is sufficient for homing pigeons (Columbia livia) to find their way home. Dennis and his colleagues developed a miniature global positioning system (GPS) tracking device, and, over a period of several weeks, trained experienced racing pigeons to fly with the devices strapped to their backs.
The birds were released in or around the Auckland Junction Magnetic Anomaly, where a cluster of massive rock slabs located 1.6 km beneath the Earth's surface causes a spatial distortion of the geomagnetic field. When their trajectories were examined (lower panel in the image below), it was found that 59 out of 92 pigeons flew up to 4km in the wrong direction before adjusting their trajectories and flying homeward. During these navigational errors, many of the pigeons flew in parallel or perpendicular to the magnetic field anomaly, for distances of between several hundred to over one thousand metres. The trajectories of others resembled a box - they flew first in parallel, and then perpendicular to the field, or vice versa. But when the pigeons flew out of the anomaly's magnetic field, they adjusted their direction, and began flying back to their lofts.
The exact nature of the navigational cues used by the pigeons is uncertain, but the researchers conclude that the pigeons used the intensity gradient - the difference in field intensity between different points in space - to determine their position. When flying in box-shaped trajectories, the birds may have been sampling the geomagnetic field information before setting off on their true course. There was no apparent correlation between the magnetic field anomaly within which the birds were released and the topographical features of the landscape, so it seems that the magnetic field was the only information used during migration.
It has been suggsted that birds (and other animals which undertake long migrations) have an internal 'compass' with which they determine their position. But it is generally believed that a compass alone is insufficient for accurate navigation; migratory birds are known to construct navigational maps, and studies of brain-damaged birds shows that the hippocampus is crucial for these spatial tasks. In the early 1950s Kramer proposed the map and compass hypothesis; according to this, the compass sets course for the destination, while the map is used to determine the bird's position relative to that destination. The Earth's magnetic field is stable over time, and contains a number of parameters which may be used by birds in constructing a navigational map. As well as the intensity gradient, the field also has a specific inclination or declination (direction). This type of information could be used to describe any point on the Earth's surface as a total field vector, consisting of the bird's position along the axes of magnetic and geographical north, as well as geographical east, west and south.
A great diversity of organisms - bacteria, insects, molluscs, crustaceans, and all five classes of vertebrates - have the ability to sense the Earth's magnetic field. However, very little is known about the neural bases of magnetoreception, and magnetoreceptors have not yet been identified. Whereas other receptors are activated by coming into physical contact with something from the external environment - e.g. photons in the case of photoreceptors, or odorant molecules in the case of olfactory receptors - this does not apply to magnetoreceptors. Because a magnetic field passes freely through biological tissues, magnetoreceptors could be located anywhere in the body.
Several lines of evidence suggest that magnetoreceptors would contain magnetite (iron oxide, Fe3O4) or greigite (iron sulphide, Fe3S4) particles. In birds, iron compound crystals thought to be magnetite have been detected in the upper parts of the beak. Structural analyses of this region of the beak show that there are clusters of these particles arranged along the insides of cell membranes at nerve terminals. These particles are superparamagnetic; i.e. they only become magnetized when exposed to an external magnetic field.
In a 2004 study by Michael Walker, who is also at the University of Auckland, pigeons were trained to jump on or off a feeding platform in response to a magnetic field. If the ophthalmic branch of the trigeminal nerve was severed, the pigeons permanently lost their ability to detect the experimenters' magnetic field. Likewise, the birds could not detect the field if a magnet was attached to the beak, or if a local anaesthetic was applied to the trigeminal nerve; in these cases, abolition of magnetoreception was reversible - when the magnet was removed, or when the effects of the anaesthetic wore off, the birds regained their ability to detect changes in the field. Furthermore, electrophysiological recordings show that neurons in the trigeminal ganglion, to which the ophthalmic branch of the trigeminal nerve projects, are responsive to changes in magnetic field intensity; these neurons are highly sensitive, responding to changes of about 0.5% of the size of the Earth's magnetic field.
Animals would require two types of magnetoreceptor - one type for detecting the direction of the magnetic field, and another for measuring the intensity of the field. According to the hypothetical models developed by Dennis and his colleagues, both types of magnetoreceptor would contain chains of magnetite crystals coupled to mechanically-gated ion channels. The receptor which detects the direction of the magnetic field would contain a chain of magnetite particles, 1.5 micrometres in length, linked by strands of microtubule-like structures to the ion channels in the receptor cells (see the image on the left in the figure below). This chain would be attached to a pivot; in response to the external magnetic field, the chain would rotate relative to a fixed plane, and align itself with the external magentic field. In the process, it would pull on one or more of the tethering strands, opening the ion channels to which they are coupled and eliciting the appropriate neural activity. Perhaps as few as six cells containing such receptors would be needed to accurately measure the direction of a magnetic field.

Hypothetical models of magentoreceptors for detecting the direction (left) and intensity (right) of thegeomagnetic field (from Walker, et al, 2002).
The receptor for detecting the intensity of the magnetic field would work in a different way. Rather than being tethered to the cell membrane, the chain of magnetite particles is free to move around in any direction over a limited range. When exposed to a magnetic field of low intensity, vibrations produced by heat energy would cause large variations in the movements of the chain, but the force of a field of greater intensity would limit the chain's movements. As in the hypothetical model for the intensity-detecting magnetoreceptor, the movements of the chain would be translated into the opening of the ion channels.
Earlier this year, Gerta Fleissner and her colleagues at the University of Frankfurt in Germany identified putative magnetoreceptors in homing pigeons. Using histological methods, electron microscopy and X-ray analysis, they visualized the distribution of magnetic iron mineral crystals in the beaks of homing pigeons, and examined their subcellular arrangement. The findings were published in the journal Naturewissenschaften.
Fleissner's group found that the crystals are located in the unmeylinated dendrite-like processes of sensory neurons in the trigeminal nerve, which project to six patches of skin lining the inner surface of the pigeon's upper beak. Each patch is approximately 350 µm long and 200 µm wide, and contains loose concentrations of crystals organized in linear or slightly curved assemblies. The system is bilaterally symmetrical, with three of these skin patches on each side of the beak.
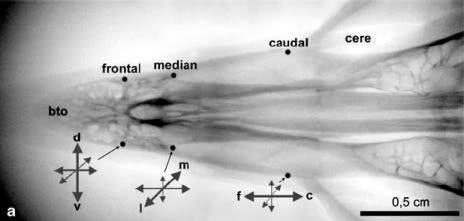
X-ray image of the upper beak showing the six areas of skin containing iron crystals and the orientation of each.
The nerve cell processes in each patch are also arranged in highly organised three-dimensional patterns: they are aligned in parallel along their longitudinal axes, with a uniform distance of about 100 µm (micrometres, thousandths of a millimetre) separating them. In each patch, the processes are oriented in a specific direction: those in the patch of skin nearest the eyes are aligned in a caudal-to-rostral (or tail-to-head) direction, those in the medial region of the beak are arranged in a middle-to-side direction, and those in the patch near the tip of the beak are arranged in a top-to-bottom direction. Each pair could, therefore, detect the one of the three axes of a magnetic field vector.
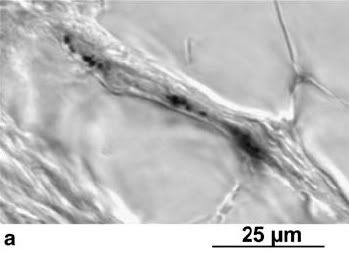
Iron crystal deposits in dendrite-like processes of the skin in the upper beak.
The processes contain two different types of iron mineral - magnetite and maghetite, organized in three subcellular compartments. The magnetite, of which about 10% of the crystals are composed, occurs as 2-4 nanometre-sized particles which are accumulated in spherical clusters of about 1 µm in diameter. These clusters are adhered to the inner surface of the cell membrane and are embedded within an organic matrix whose composition is as yet unknown. By contrast, the magnetite crystals are found as an irregular crust on the surface of vesicle-like structures, or in flat, square platelets that form approximately 10 µm-long chains along the length of the processes. The authors estimate that the homing pigeon's beak contains about 85 nanograms of iron.
The mechanism by which this system transduces magnetic signals remains unclear however, although the authors suggest that the structures they have identified could detect both the strength and orientation of the geomagnetic field. They speculate that the maghetite crystals around the vesicle-like structures may be involved in amplifying the intensity of a local magnetic field. This would account for the high degree of sensitivity required by a magnetoreceptor. Because they have found iron oxide crystals in the beaks of robins, golden warblers and chickens, the authors also suggest that this system may be a universal found in all birds.
Much work remains to be done before we gain any real understanding of magnetoreception and the construction of navigational maps by birds and other migratory animals. Although birds are amenable to behavioural experiments, they are not particularly conducive to neurobiological studies. Recently, however, it was discovered that fruit flies and zebrafish are sensitive to magnetic fields. These are among the favourite model organisms of neurobiologists, and the discovery that they too can detect magnetic fields should allow researchers to investigate magnetoreceptive mechanisms in greater detail. Also, a better understanding of navigational cues should be facilitated by the continuing development of GPS technologies, which will enable researchers to gather high resolution data from migrating birds.
References:
Dennis, T. E, et al. (2007). Evidence that pigeons orient to geomagnetic intensity during homing. Proc. R. Soc. B. DOI: 10.1098/rspb.2007.3768. [Full text]
Fleissner, G., et al. (2007). A novel concept of Fe-mineral-based magnetoreception: histological and physicochemical data from the upper beak of homing pigeons. Naturewissenschaften DOI: 10.1007/s00114-007-0236-0. [Full text]
Johnsen, S. & Lohmann, K. J. (2005). The physics and neurobiology of magnetoreception. Nat. Rev. Neurosci. 6:703-712. [Full text]
Mora, M. V., et al. (2004). Magnetoreception and its trigeminal mediation in the homing pigeon. Nature 432: 508-511. [Abstract]
Walker, M. M. et al. (2002). The magnetic sense and its use in long-distance navigation by animals. Curr. Op. Neurobiol. 12: 735-744. [Full text]
Gagliardo, A., et al. (1999). Homing in pigeons: The role of the hippocampal formation in the representation of landmarks used for navigation. J. Neurosci. 19: 311-315. [Full text]
Related:
Very awesome! I think navigating (and orienteering) in birds, and other creatures is fascinating. I'm also continually impressed with the methods biologists have used to examine these phenomena. The smell-based navigation experiments in pigeons at the University of Pisa I think are well thought out and measured. The most entertaining that I've read recently however is this article where biologists hypothesized that ants have an internal pedometer to measure distance from home, and to test the theory they put ants on stilts (and cut others legs in half).
A while back, noted pseudoscientific babbler Rupert Sheldrake wrote a "Dangerous Ideas" essay in which he said that animals' ability to navigate long-distance migrations suggested that they had "a sense of direction involving new scientific principles" (no doubt his "morphic field"). I took the time to complain about this, as part of my general dissatisfaction with the "Dangerous Ideas" thing, so I'm particularly pleased to see such details.