

Unicolonial ants, such as these Argentine ants (Linepithema humile), are genetically unrelated
but will cooperate to defeat a much larger adversary.
Source: Alex Wild / Live Science
 It has been a mainstay of evolutionary theory since the 1970s. Natural selection acts purely on the level of the individual and any cooperation observed between organisms merely hides a selfish genetic motive. There have been two pioneering theories to explain cooperation in the natural world given this framework: the first was William Hamilton's (1964) theory of kin selection and the second was Robert Trivers' (1971) theory of reciprocal altruism.
It has been a mainstay of evolutionary theory since the 1970s. Natural selection acts purely on the level of the individual and any cooperation observed between organisms merely hides a selfish genetic motive. There have been two pioneering theories to explain cooperation in the natural world given this framework: the first was William Hamilton's (1964) theory of kin selection and the second was Robert Trivers' (1971) theory of reciprocal altruism.
However, both of these scenarios break down where it comes to unicolonial ants. In a paper in the journal Trends in Ecology and Evolution (subscription required) Heikki Helantera, of the University of Sussex, and colleagues at Rice University have investigated how previous theories to explain cooperation don't apply for these unique supercolonies.
Unicolonial ants carry polydomy [multiple nests in a supercolony that all individuals rotate through] and polygyny [multiple queens in one nest] to extremes. Colonies are huge, each being a network of hundreds or thousands of nests, each with multiple queens. There is no worker aggression, and there is free movement among nests on a vast scale. The energy that might have been put into fighting and territoriality flows into the common good, more ants.
Such a concept, a form of genuine anarchism in the animal world, was thought to be impossible given existing theory. These ants live in colonies where relatives exist but, with so much migration throughout a network stretching thousands of kilometers, each ant worker is mostly surrounded by total strangers that share none of their genes. Only one other species has ever been known to organize themselves in such a fashion (and if you're reading these words right now you know who you are).
To understand how unicolonial ants have come to be the way they are, we must first understand what they're not. Kin selection has proposed that cooperation will emerge in groups that are made up of close relatives. Hamilton's rule, beautiful in its simplicity, proposed that cooperation occurs when the cost to the actor (C) is less than then the benefit to the recipient (B) multiplied by the genetic relatedness between the two (r). This equation is written out simply as rB > C.

Lion siblings often cooperate as teams and benefit through kin selection.
Source: Scotch Macaskill / Wildlife Pictures.com
To put this into context: an alpha male lion and his brother share half of their genes, so have a genetic relatedness of 0.5. Suppose this brother recognizes that the alpha male is getting old and could easily be taken down. If so, the brother could potentially have eight additional cubs (just to pull out an arbitrary number). But, instead, that brother decides to help the alpha male to maintain his position in the pride and, as a result, the alpha ends up having the eight additional cubs himself while the brother only has five. The brother has lost out on 3 potential cubs. But, even so, because he assisted his brother he has still maximized his overall reproductive success from a genetic point of view: (0.5) x 8 = 4 > 3. He could have attempted to usurp his brother and, perhaps, had the eight cubs himself but he wouldn't have been in any better of a position as far as his genes were concerned.
Reciprocal altruism follows this same basic idea, but proposes a mechanism that could work for individuals that are unrelated. In this scenario, cooperation occurs when the cost to the actor (C) is less than the benefit to the recipient (B) multiplied by the likelihood that the cooperation will be returned (w) or wB > C. This has been demonstrated among vampire bats who regurgitate blood into a stranger's mouth if they weren't able to feed that night. Previous experience has shown the actor that they're likely to get repaid if they ever go hungry one night themselves. This theory requires that individuals be part of a single group, with low levels of immigration and emigration, so that group members will be likely to encounter each other on a regular basis.
Previously, it was argued that all ants followed an extreme form of kin selection. Because of their unique process of reproduction females develop from fertilized eggs and have paired chromosomes (that is, one from each parent). However, males develop from unfertilized eggs and only have a single chromosome from their mother. As a result, female workers share up to 75 percent of their genes with sisters but only 50 percent with their mother (or their own offspring, if they were to reproduce). Worker ants therefore have greater genetic success by not reproducing but, instead, helping to raise and protect their legion of closely related sisters.
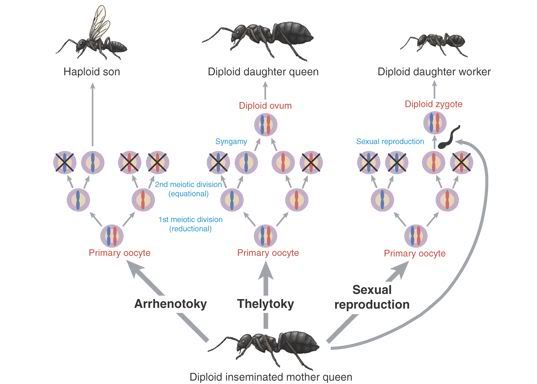
Ant reproduction gives rise to genetic sisterhood.
Source: Unattributed
This explanation has been somewhat clouded given more recent evidence that queens engage in polyandry (mating with multiple males). A queen will frequently mate with up to five different males and store their combined sperm, around 100 million of them, in a special compartment called the spermatheca. By releasing a single sperm at a time the queen can control the number of eggs she lays. However, because there are multiple fathers, the genetic relationship between the female worker ants is reduced. Female workers may therefore only be related by 25 percent with the females they're helping to raise. Why would female workers continue to be non-reproductive and help rear distant relatives when they could have twice the reproductive success by having their own offspring? While there are strategies female workers employ to maximize their own reproductive success (like preferrentially rearing eggs that they are more closely related to or, in some rare cases, reproducing themselves) it still remains puzzling why ants have been so successful given this seeming contradiction.
If you add to this the realities of multiple queens in a single nest (polygyny) and supercolonies that are composed of thousands of such nests (polydomy), the problem becomes insurmountable. If worker ants share zero percent of their genes with those they're cooperating with, as is the case in these unicolonies, then why cooperate? What do they have to gain?
This is the problem that Helantera and colleagues are seeking to understand in their latest paper. While the authors emphasize a range of possible explanations, I want to focus on just one that has been generating a great deal of interest in the last few years: group selection.
The extreme cooperation of unicolonial ants has been suggested to be an example of selection occurring on levels higher than the individual, such as the superorganism, group or even population.
Group selection is the idea that, under certain circumstances, genes will be selected for because they benefit the overall success of the group rather then simply the individual. While it is usually assumed that these populations will have a high level of relatedness (making the promotion of the group an extended form of kin selection) the authors suggest a scenario in which group selection could apply even among unrelated group members.

{kind=link}
This is a possibility I like to call Ronald Reagan's Alien Invasion Hypothesis. In a speech before the United Nations on Sept. 21, 1987 Reagan stated that:
In our obsession with antagonisms of the moment, we often forget how much unites all the members of humanity. Perhaps we need some outside, universal threat to make us recognize this common bond. I occasionally think how quickly our differences worldwide would vanish if we were facing an alien threat from outside this world.
So under this possibility a common threat to all colony members would outweigh the low level of genetic similarity because, unless everyone pulls together, the entire group is in jeopardy. If one colony was competing with a rival colony then selection for individual selfishness could drive the population to extinction while selection for cooperation would allow the colony to thrive.
Under this view, extant unicolonial populations are the ones that have not yet succumbed to selfishness. Relatedness and mutual policing select against selfishness in non-unicolonial populations, but stop applying when relatedness decreases to zero. . . [However], constraints arising from the natural history of the species or pleiotropic effects of selfish genes, might prevent selfish genotypes from arising even under zero relatedness.
This cooperation could then continue long after the initial threat was gone under the force of phylogenetic inertia. Perhaps, in the future, selection would cause the unicolony to break into smaller, more genetically similar colonies once the impetus for group selection no longer exists? Or perhaps the benefits of cooperating with strangers simply outweighs the costs of competition and natural selection has produced a genuinely altruistic society?

Unicolonial cooperation has inspired activist art such as this print from the Beehive Collective.
Source: Beehive Collective
At the current time there are 31 known unicolonial ant populations around the globe. This is a small minority given the more than 12,000 described species. However, given that research on unicolonial ants is so new, there is still a great deal of research that needs to take place concerning this unique experiment of the natural world. At the very least, unicolonies provide us with a source of inspiration and the ability to marvel at the amazing beauty and diversity of the natural world. With the knowledge that stable supercolonies composed of strangers continue to thrive in nature, perhaps there's something we could learn from those creatures that first invented this approach.
References:
Hamilton, W.D. (1964). The genetical evolution of social behaviour I and II. -- Journal of Theoretical Biology 7: 1-16 and 17-52
Trivers, R.L. (1971). The evolution of reciprocal altruism. Quarterly Review of Biology. 46: 35-57.
Helantera, H., Strassman, J.E., Carrillo, J., Queller, D.C. (2009). Unicolonial ants: where do they come from, what are they and where are they going? Trends in Ecology and Evolution. doi:10.1016/j.tree.s009.01.013
Nice summary. A couple of thoughts.
Inbreeding is potentially important. As always.
I think it is important to distinguish betweeen "superorganism" and "group selection." Just in case.
Finally, if there are fewer than 100 species of ant that is over the top in their behavior in two areas (multi-lineality and eusociality) at the same time, that means a very very small percentage of this sort of creature has a behavior syndrome that is explained primarily by phylogeny and not adaptively.
Obviously, these unicolonial ants proves selection occurs at the group level, and we can finally settle the probelm of unit of selection once and for all.
/snark toward the evolutionary biology community
When theorizing why the high level of cooperation between unrelated ants, how do you distinguish between the truth and a "just-so" story?
It seems like a sustainable strategy such as Tit for Tat could explain this apparent altruism. In smaller colonies, the Tit for Tat strategy becomes prominent due to its sustainability. Then, when ants begin for one reason or another to group together into larger and larger colonies, the Tit for Tat mentality/gene/whatever becomes impossible to invade and so cooperation continues indefinitely.
If you have not already noticed, ants are evolving!
So Reagan had read Watchmen?
Erm, forgive me if I'm missing something here but how is this any different from a mutually beneficial relationship [symbiosis]? This species of ant gains little from competing among its own species [the rare method of queen-production negates the most common form of inter-species competition: for mates]. In your words: "the benefits of cooperating with strangers simply outweighs the costs of competition and natural selection [and] has produced a genuinely altruistic society" though with two caveats.
First, I think it's inaccurate to claim natural selection itself has a "cost". The actions associated with it [competing, mating, rearing the young] do have costs in time and energy but it's not accurate to say Naturally Selecting "costs" more than Not Naturally Selecting.
Second, It is not in fact "altruistic" as that would imply not acting in the best interests of ones genes. The worker ants as you have said are of course not the actual reproducers. They function much more like an Extended Phenotype [/DawkinsWorship.exe<3] I realize we usually think of Symbiosis in terms of inter-species relationships but otherwise this is the textbook definition of Mutualism.
And now to destroy all my pretensions of intellectualism with some fanfare:
=p w00t, Selfish Gene 4 LYFE.
That's not the selfish gene theory, as it was explained to me...
As I understood it, the selfish gene was the idea that each gene "tried" to propagate itself as far as possible. If that can be done by creating animals with an almost perfect altruism for any creature they can breed with, then that is a valid path for a gene's evolution to take. I'm sure that any of these colonies acting selfishly would trigger something that would drive the other colonies in the supercolony to attack it. Once established, mutual altruism is pretty much self-supporting.
Look at it this way: Each ant colony is one breeder and a set of more-or-less clones that rely on the breeder for the propagation of their genes, that is, they can be viewed as a single individual. If this "individual" is in, say, a small valley with one other "individual" and they fight, they're both going to do a lot worse than if they were in the valley alone. But if some mutation, or perhaps a close relationship between the two colonies, causes them to work together, they can ignore each other and concentrate on more important threats. They're getting a concrete benefit out of it, which disappears if they behave selfishly on a smaller scale. Cooperation is ultimately selfish, because rather than making the best of the environment you have, it changes the environment so that even a small slice is far better than what you had before.
If very simplistic terms: What if there is a gene for cooperation? wouldn't that just give further credence to the selfish gene theory?
As wazza said, your definition of selfish gene theory is a little unusual, and it is not consistent with what you say here:
Group selection is the idea that, under certain circumstances, genes will be selected for because they benefit the overall success of the group rather then simply the individual.
So this would not actually conflict with selfish gene idea, it's just that in this case the "selfish genes" propagate by benefiting the group at the expense of the individual.
Polyfidelity would be very helpful for humans to have group families with fewer total children shared for a no growth sustainable world with biodiversity of intimany and much more stability and security from mate loss of all kinds. It would allow pooling of resources and more homekeeping and nurturing in a complex eco-economically stressed world of the 21st century. As opposed to the toxic nuclear family of over-consumption and isolation in padded suburban pill-boxes of insecurity where we now have 2 full-time worker barely making the ends meet.
Ah, excuse me, Comment #2? Comment #1 already inoculated this thread against you. Comment #3? We already have our just-so story in plae, thank you.
But seriously, comment #3: The best way to do this besides replication (of results, not ants) is to prove a cost. Adaptations that have a cost are more likely to be real. That is true to the extent that many of us consider "adaptations" that don't have a cost to not be real adaptation (though in theory such things are possible).
#9: The gene for cooperation is the hypothetical construct that works together with kin selection given haplodiploid reproduction and the relatedness of sisters at 0.75 , which goes away wiht polyandry and multiple queens.
#10: Right. Group selection is simply a level of selection argument whereby a group is a unit of replication. It works perfectly if you think of the cells in a metazoan as being a group of cells because they share specialized replicators. It goes away as a theoretical possibility as soon as all the individuals can follow separate strategies, and it is impossible or almost impossible to make it come back.
I never have been able to take the selfish gene theory(in these specific applications seriously)
Why couldn't all the ants have genes for general altruism?
How does it go from protein to altrusim?
What feedback is there?
How clear in the intergrity of this feedback?
Can the organisms identify alleles FOR altruism? Or do they just practice altruism to those near them
Can it even be said that there is an allele FOR altrusim or
any identidied gene network that causes this?
What about the occasional game theory cheaters? wouldn't they blow thorough such a system and exploit, making this not an evolutionary stable strategedy
Are these unicolonial species monophyletic? Do we have any idea of how old this behavior is? How much genetic diversity is there within each of these species? Could this perhaps be the result of kin selection gone awry after a bottleneck event?
Perhaps it's not an ESS but a cheater hasn't appeared yet, or perhaps cheating is just too costly.
One problem with this article is that unicolonial Argentine ants are actually very related to one another (in contrast to what is said above). In fact, workers share many more genes than would be expected for such a polygynous society. The high levels of relatedness among workers within colonies of introduced populations of Argentine ants (compared to other species or even nests of other Argentine ants in their native range) suggest that kin selection theory is sufficient to explain unicoloniality and group selection does not have to be invoked.
@13: The trouble with a generalized "gene" for altruism (it would be more appropriate to refer to it as a gene network) is for the very reason you bring up: cheaters and free riders. Therefore the selfish gene theory predicts altruism would only be found in closely related individuals (kin selection) or in defined groups where an altruistic act can be paid back (reciprocal altruism). Unicolonial ants are neither closely related nor in defined groups. Other approaches, such as generalized reciprocity or the "genial gene" theory of Joan Roughgarden, propose alternative frameworks. Clearly, further research is necessary to create a unified framework of cooperative behavior.
And everything Greg said.
Another thing. Everyone forgets the "b" in Hamilton's equation. Kin selection can work under very low values of relatedness if "b" is high enough. I imagine if you are very ecologically successful (living at extremely high densities and reproducing wildly) - as many unicolonial ants do - r can be quite low and still fit the model.
Our (humanity's) ideas on selection and evolution have evolved from Darwin, to Hamilton, and now to Helantera's notion of "group selection". Fascinating how our ideas evolve. Maybe these unrelated ants help each other out because the Queen ants mate with multiple males, and so the males do not know what offspring is theirs; or maybe they act the same way that the bats do, which is "I help you, and you help me when I need it", etc. This paper tells us that the "stable supercolonies" benefit all the individuals in the group. Some benefits are: prevents inbreeding, so there are better chances of stronger/better genes being passed on (âshuffling of the deckâ); by helping one another, competition/death is avoided. According to this paper, the goal of the supercolonies is to produce more ants. There is a better chance of reproduction if these ants stick to "group selection" in order to produce a larger population. Humans also have group selection as well. We help those who are unrelated because maybe our genes/offspring are mixed within our groups. If we protect our group, we are therefore protecting our genes (just like these ants). I think this "group selection" is a subcategory of Hamilton's "kin selection", or vice versa.
Nice to see our work raises a discussion!
As the author of the paper, some comments:
1. megan hoy is right: group selection and kin selection are two different mathematical approaches to modelling natural selection, but if the models are built correctly, produce the same results. Since social insects live in family groups, I find a "Hamiltonian" kin selection or inclusive fitness terminology and approach much more useful, and prefer to use kin selection terminology, as do most researchers in the field.
quoting myself in the paper:
"Thus, the superorganism view does not change the argument that adaptations that benefit the group instead of the individual are only predicted to arise and be maintained under positive relatedness, when individuals act to maximize their inclusive fitness. This Hamiltonian kin-selection conclusion is not changed by considering selection above the level of the individual."
and again (this sentence immediately follows the last quote form our paper in the blog above):
"This might affect the distribution of traits in extant unicolonial populations, even if population-level selection per se cannot lead to unicoloniality."
2. In the paper we describe how supercolonies are the result of kin selected behavioural rules, but when colonies grow to extreme sizes (i.e. when a supercolony covers the whole breeding population, especially in invasive populations), the same rules are no longer increasing the inclusive fitness of the workers. However, the workers are unable to assess the situation, and behave according to their now maladaptive rules. Like Amplexus suggests above, the workers pratice altruism towards those near them, because during their evolutionary history these individuals (nestmates) have been relatives.
3. We make the point in our paper that supercolonies seem to start out as family units, but when they grow to extreme sizes, relatedness approaches zero. This is because relatedness in the technical sense of kin selection theory/inclusive fitness theory is measured as the genetic similarity of individuals compared to random individuals in the population. Relatedness is always measured relative to to a background population, and changing the background population changes relatedness. If the whole population, the area over which competition for genetic representation in the future population occurs, is covered by the same family, and individuals mix across nests, immediate nestmates are not more related than random individuals. According to the same logic, as soon as there is any competition between e.g. two neighbouring supercolonies, relatedness within a supercolony increases above zero, because the scale of measuring relatedness changes.
4. In the long term, the huge supercolonies are predicted to be evolutionary dead ends. If relatedness is zero, selection does not affect worker traits, and also selfish behaviour is not selected against. Unicolonial lineages seem to be dead-ends in the light of the available phylogenetic data. More data is obviously needed, but the current evidence supports the view that supercolonies do not thrive in the very long run. As predicted by a Hamiltonian viewpoint.
A link to an open access paper that covers partly the same issues http://jbiol.com/content/pdf/jbiol154.pdf
Best regards,
Heikki Helanterä
Sometimes the sheer audacity of the lies by evolutionists is breathtaking. This example is a case in point.
Creationists (and non-creationists too, for that matter) have pointed out many times over the years that modern racism relies on evolutionary thinking for justification, and that the Nazi ideology of an Aryan Master Race comes directly out of an evolutionary mindset. This is pretty straighforward and I won't argue it further here, but naturally the more extreme evolutionary types don't like having these sorts of things pointed out. Among creationists, many of the best and most well-researched articles documenting and exposing evolutionary racism and Nazi-evolution connections have been written by...
Jerry Bergman.
This claim about Dr. Bergman completely turns reality on its' head. Since when does fighting racism make you a racist? If you look at the words, that is exactly what Dr. Bergman was accused of by Mr. Flank. While some may try to justify it as "OK" for one reason or another, employment of racial quotas and other tactics boils down to simply that: racism against whites. The rule "two wrongs do not make a right" has apparently been forgotten.
As this poster mentioned, Dr. Bergman has also written for Destiny, which is a superb magazine operated by and for black Americans! And that is just the tip of the iceberg when it comes to his exposing the lies and hypocrisy of evolutionists. Here is a list of a few other articles he has authored fighting racism and fascism (with thanks to CRSnet participants for some of these):
Bergman, Jerry, "A Brief History of the Eugenics Movement" CEN Tech. J., vol. 5(2), 1991, pp. 143-153
ABSTRACT
Eugenics, the science of improving the human race by scientific control of breeding, was viewed by a large segment of scientists for almost one hundred years as an important, if not a major means of producing paradise on earth. These scientists concluded that many human traits were genetic, and that persons who came from genetically 'good families' tended to turn out far better than those who came from poor families. The next step was to encourage the good families to have more children, and the poor families to have few or no children. From these simple observations developed one of the mostfar-reaching movements, which culminated in the loss of millions of lives. It discouraged aiding the sick, building asylums for the insane, or even aiding the poor and all those who were believed to be in some way 'genetically inferior', which included persons afflicted with an extremely wide variety of unrelated physical and even psychological maladies. Their end goal was to save society from the 'evolutionary inferior'. The means was sexual sterilization, permanent custody of 'defective' adults by the state, marriage restrictions, and even the elimination of the unfit through means which ranged from refusal to help them to outright killing. This movement probably had a greater adverse influence upon society than virtually any other that developed from a scientific theory in modern times. It culminated with the infamous Holocaust and afterward rapidly declined.
Bergman, Jerry, "Eugenics and the Development of Nazi Race Policy," PERSPECTIVES ON SCIENCE & CHRISTIAN FAITH Vol 44, no. 2, June 1992.
ABSTRACT
A central government policy of the Hitler administration was the breeding of a "superior race." This required, at the very least, preventing the "inferior races" from mixing with "superior" ones in order to reduce contamination of the latter's gene pool. The "superior race" belief is based on the theory of group inequality within each species, a major presumption and requirement of Darwin's original "survival of the fittest" theory. A review of the writings of Hitler and contemporary German biologists finds that Darwin's theory and writings had a major influence upon Nazi policies. Hitler believed that the human gene pool could be improved by selective breeding, using the same techniques that farmers used to breed a superior strain of cattle. In the formulation of his racial policies, he relied heavily upon the Darwinian evolution model, especially the elaborations by Spencer and Haeckel. They culminated in the "final solution," the extermination of approximately six million Jews and four million other people who belonged to what German scientists judged were "inferior races."
Bergman, Jerry "Evolution and the Development of Nazi Race Policy," Contrast (Bible Science Newsletter insert), Nov-Dec 1988, vol 7., No. 6.
Links Hitler's interpretation of "the survival of the fittest" doctrine to the Nazi holocaust.
Bergman, Jerry, "Evolution and the Origins of the Biological Race Theory," Creation Ex Nihilo Technical Journal 7(2):162-168.
ABSTRACT
Scientists have almost universally concluded that the human races are largely equal in regard to innate intelligence and most other traits. In spite of the wide social disparities between the races in the West, no more than approximately one standard deviation difference in mean intelligence exists between the whites and blacks. This fact is contrary to a basic requirement of naturalistic evolution: in order for selection to take place, differences must exist between individual organisms for selection to select from. For selection to work something first must cause races to develop, a process which in Darwinian terms is called speciation. As evolution progresses, the contrast between groups must become greater, producing development of new definable divisions. The lack of major differences between races, especially -in intelligence, the factor most crucial for the major contrast between Homo sapiens and 'lower' forms of life, creates a major difficulty for current evolution theory. In addition, misuse of the theory of evolution was an important factor in the extreme forms of racism, especially that against blacks and Jews, that flourished at the turn of the century and for many years beyond.
Bergman, Jerry "Evolution, Race, and Equality of Intelligence," Creation Research Society Quarterly, Sept. 1980, 17:127-134.
Emphasizes lack of important differences among "races," and points the finger at evolution for the cause behind prejudices against blacks and Jews at the turn of the 20th cent.
Bergman, Jerry, "Ota Benga: The man who was put on display in the zoo!," Creation Ex Nihilo 16(1):48-50.
"What would you do if you went to the zoo and saw a black African on display in a cage? It happened this century! Read about this tragic case of evolutionary racism at its worst."
Bergman, Jerry, "NINETEENTH CENTURY DARWINISM AND THE TASMANIAN GENOCIDE," Creation Research Society Quarterly, (Vol.??) Received 19 September 1994; Revised 29 April 1995
Abstract
It was widely believed in the nineteenth century that the Tasmanians were a living link between modern humans and their primate ancestors. Given the presupposition of naturalistic evolution, the Tasmanian people were often seen as less than human and, consequently, many people felt that it was not wrong or immoral to treat them like animals. This attitude eventually influenced behavior which resulted in the total extermination of the native Tasmanians. Today it is concluded that they were a distinct racial group similar to the Australian Aborigines that possessed a unique culture and were fully human. This event is only one of many examples of the numerous tragedies that evolutionary naturalism has produced in modern times.
Bergman, Jerry, "The Teaching of Hate: The Influence of Biological Evolution on Modern Literature," Creation Ex Nihilo Technical Journal 6(2):185-193.
I've met Dr. Bergman and find him to be a quiet, humble servant of Christ, and as erudite as anyone I've ever met. That evolutionists are reduced to such scurrilous lies in fighting science tells me something about the bankruptcy of their religion.
J.W. adds (by permission):
I talked with Jerry Bergman on the phone over the weekend. Fact of the matter is that Jerry NEVER gave money to any racist group. He DID write letters to racist groups in order to solicit their basis for their racism. This was part of his research on evolution and racism. One of his letters WAS published in a racist publication. But then again, some of his letters have been published in atheistic publications as well.
Finally, Jerry said that he has heard similar insinuations for some ten years now, and remarked that, if that is the best the anti-Creationists can do against him, they really have nothing to stand on!
The "unattributed" image is from Gadagkar (2004), "Sex⦠only if really necessary in a feminine monarchy." Science 306 (5702): 1694-1695.